Dengue virus
Dengue virus (DENV) is the cause of dengue fever. It is a mosquito-borne, single positive-stranded RNA virus of the family Flaviviridae; genus Flavivirus.[1][2] Four serotypes of the virus have been found, a reported fifth has yet to be confirmed,[3][4][5] all of which can cause the full spectrum of disease.[1] Nevertheless, scientists' understanding of dengue virus may be simplistic as, rather than distinct antigenic groups, a continuum appears to exist.[6] This same study identified 47 strains of dengue virus.[7] Additionally, coinfection with and lack of rapid tests for zika virus and chikungunya complicate matters in real-world infections.[8]
| Dengue virus | |
|---|---|
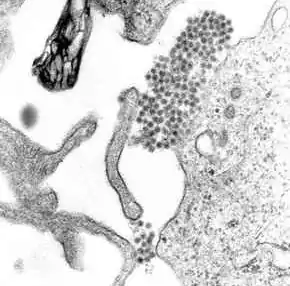 | |
| A TEM micrograph showing dengue virus virions (the cluster of dark dots near the center) | |
| Virus classification | |
| (unranked): | Virus |
| Realm: | Riboviria |
| Kingdom: | Orthornavirae |
| Phylum: | Kitrinoviricota |
| Class: | Flasuviricetes |
| Order: | Amarillovirales |
| Family: | Flaviviridae |
| Genus: | Flavivirus |
| Species: | Dengue virus |
Dengue virus has increased dramatically within the last 20 years, becoming one of the worst mosquito-borne human pathogens which tropical countries have to deal with. Current estimates indicate that as many as 390 million infections occur each year, and many dengue infections are increasingly understood to be asymptomatic or subclinical.[9]
Evolution
Based on the analysis of the envelope protein, at least three genotypes (1 to 3) are known. In 2013, a fourth serotype was reported.[3] A single report of a fifth serotype DEN-5 in 2015[10] has not been replicated or further reported on.[5] The rate of nucleotide substitution for this virus has been estimated to be 6.5×10−4 per nucleotide per year, a rate similar to other RNA viruses. The American African genotype has been estimated to have evolved between 1907 and 1949. This period includes World War I and World War II, which were associated with considerable movement of populations and environmental disturbance, factors known to promote the evolution of new vector-borne viral species.
A Bayesian analysis of all four serotypes estimated that their most recent common ancestor existed about 340 AD (95% confidence interval: 280 BC-850 AD).[11]
Lifecycle
Until a few hundred years ago, dengue virus was transmitted in sylvatic cycles in Africa, Southeast Asia and South Asia between mosquitoes of the genus Aedes and nonhuman primates, with rare emergences into human populations.[12][13] The global spread of dengue virus, however, has followed its emergence from sylvatic cycles and the primary lifecycle now exclusively involves transmission between humans and Aedes mosquitoes.[14] Vertical transmission from mosquito to mosquito has also been observed in some vector species.[15] Dogs have been found to be infected by the virus, but more research is needed to determine if dogs or other animals can serve as reservoirs or are just incidental hosts.[16]
Recent findings suggest that as the virus infects human cells, host homeostatic processes such as autophagy and ER stress response, not to mention apoptosis, are triggered depending on the infected cell type.[17] The activation of autophagy and ER stress during infection enhances virus reproduction.[18] [19] Attempts to provide detailed summaries of the life cycle of dengue at the cellular level are published in review articles from different research groups.[20] [21]
Genome
The DENV genome is about 11000 bases of positive-sense, single stranded RNA (ssRNA) that codes for three structural proteins (capsid protein C, membrane protein M, envelope protein E) and seven nonstructural proteins (NS1, NS2a, NS2b, NS3, NS4a, NS4b, NS5).[4] It also includes short noncoding regions on both the 5' and 3' ends.[1][22]
Structural and enzymatic proteins
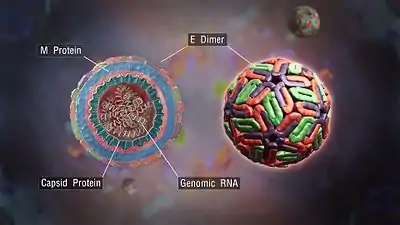
E protein
The DENV E (envelope) protein, found as a dimer on the surface of the mature viral particle, is important in the initial attachment of this particle to the host cell. Each E protein monomer comprises three ectodomains, ED1 to ED3, and a transmembrane segment. ED2 includes the dimerization interface, two glycosylation sites, and the peptide of fusion with the cellular membrane. ED3 is a continuous polypeptide segment; its fold is compact and immunoglobulin-like.[23][24] Dengue virus is transmitted by species of the mosquito genus Aedes. Several molecules that interact with the viral E protein (ICAM3-grabbing nonintegrin, CD209,[26] Rab 5,[27] GRP 78,[28] and the mannose receptor[29]) have been shown to be important factors mediating attachment and viral entry.[24] The membrane form of ribosomal protein SA may also be involved in the attachment.[30] E protein is known to contain physicochemically conserved B cells and T cells specific epitopes, which can be exploited to design vaccine.[31] Recombinant domains of the E protein are used as well-defined antigens in the serological detection of antibodies directed against dengue virus and as immunogens in vaccine candidates.[32][33][34]
prM/M protein
The DENV prM (membrane) protein, which is important in the formation and maturation of the viral particle, consists of seven antiparallel β-strands stabilized by three disulfide bonds.[24]
The glycoprotein shell of the mature DENV virion consists of 180 copies each of the E and M proteins. The immature virion starts out with the E and prM proteins forming 90 heterodimers that give a spiky exterior to the viral particle. This immature viral particle buds into the endoplasmic reticulum and eventually travels via the secretory pathway to the Golgi apparatus. As the virion passes through the trans-Golgi network, it is exposed to low pH. This acidic environment causes a conformational change in the E protein, which disassociates it from the prM protein and causes it to form E homodimers, which lie flat against the viral surface, giving the maturing virion a smooth appearance. During this maturation, pr peptide is cleaved from the M peptide by the host protease, furin. The M protein then acts as a transmembrane protein under the E-protein shell of the mature virion. The pr peptide stays associated with the E protein until the viral particle is released into the extracellular environment. This pr peptide acts like a cap, covering the hydrophobic fusion loop of the E protein until the viral particle has exited the cell.[24]
NS3 protein
The DENV NS3 is a serine protease, as well as an RNA helicase and RTPase/NTPase. The protease domain consists of six β-strands arranged into two β-barrels formed by residues 1–180 of the protein. The catalytic triad (His-51, Asp-75 and Ser-135) is found between these two β-barrels, and activity is dependent on the presence of a 43 amino acid segment of the NS2B cofactor. This cofactor wraps around the NS3 protease domain and becomes part of the active site. The remaining NS2B residues before and after the cofactor region contain helical domains involved in membrane binding. The remaining NS3 residues (180–618) form the three subdomains of the DENV helicase. A six-stranded parallel β-sheet surrounded by four α-helices makes up subdomains I and II, and subdomain III is composed of four α-helices surrounded by three shorter α-helices and two antiparallel β-strands.[24]
NS4A protein
DENV NS4A is a nonstructural protein involved in altering cell membrane curvature[35] and induction of autophagy.[19] In addition to its membrane altering property, NS4A is a scaffold for the virus replication complex and undergoes oligomerization.[36] Mutations of NS4A that affect interaction with NS4B abolished or severely reduced virus replication indicating the importance of NS4A and its interaction with NS4B in dengue reproduction.[37]
NS5 protein
The DENV NS5 protein is a 900-residue peptide with a methyltransferase domain at its N-terminal end (residues 1–296) and a RNA-dependent RNA polymerase (RdRp) at its C-terminal end (residues 320–900). The methyltransferase domain consists of an α/β/β sandwich flanked by N-and C-terminal subdomains. The DENV RdRp is similar to other RdRps containing palm, finger, and thumb subdomains and a GDD motif for incorporating nucleotides.[24]
Complexes between the E protein and neutralizing antibodies
Crystal structures of complexes between antibodies and either the ectodomain (sE) of the viral E protein or its domain 3 (ED3) have helped understand the molecular bases of the virus recognition and neutralization. Some of the epitopes are partially or totally inaccessible in the known structure of the mature virion. The corresponding antibodies are, therefore, assumed to bind to alternate or transitional conformations of the virus at 37 °C.
- The murine antibody E111 neutralizes DENV1. Its Fab and scFv fragments were crystallized in complex with the ED3 domain of DENV1. Its epitope is located around β-strands C and C' of ED3, and the intervening loop.[38]
- The murine antibody 1A1D-2 strongly neutralizes DENV1, DENV2, and DENV3. Its Fab fragment was crystallized in complex with the ED3 domain of DENV2. Its epitope straddles β-strands A and G of ED3.[39]
- The murine antibody 2H12 cross-reacts with all four DENV serotypes. It neutralizes the corresponding viruses, except DENV2. Its Fab fragment was crystallized in complex with the ED3 domains of DENV1, DENV3 and DENV4. Its epitope is located around the conserved AB loop of ED3.[40]
- The murine antibody 4E11 neutralizes all four DENV serotypes with varying efficacies. Its scFv fragment was crystallized in complex with the ED3 domains of the four DENV serotypes. Its epitope straddles β-strands A and G of ED3 as does the epitope of 1A1D-2.[41][42] The structures at 2.0 Å resolution have enabled one to analyze the roles of water molecules within the protein interfaces and the roles of somatic hypermutations outside of these interfaces in the interactions and cross-recognitions.[43]
- The chimpanzee antibody 5H2 potently neutralizes DENV4. Its Fab fragment was crystallized in complex with the sE protein of DENV4. Its epitope is included in domain 1 (ED1) of the E protein.[44]
- The human antibodies Ede1-C10, Ede2-A11, and Ede2-B7 potently neutralize all four DENV serotypes. Their Fab or scFv fragments were crystallized in complex with the sE protein of DENV2. The recognition determinants of these antibodies are at a serotype-invariant site in the E dimer interface and include the exposed side chains of the E fusion loop and the two conserved glycan side-chains.[45]
Disease
Common names for dengue fever include breakbone fever, vomiting and dandy fever; dengue hemorrhagic fever and dengue shock syndrome are the severe forms.[46] Dengue is found in tropical and subtropical climates worldwide, mostly in urban and semiurban areas.[47] People of all ages who are exposed to infected mosquitoes are at risk for developing dengue fever. The disease occurs most often during the rainy season in tropical countries in Southeast Asia, South Asia and South America, with high numbers of infected mosquitoes.[48] The virus is transmitted to humans through the bites of infected female mosquitoes, though humans are not capable of transmitting the disease and are not contagious.[47][49][48] The incubation period is 3 to 14 days, while the period of the illness is 3–7 days.[49][50] Signs and symptoms may include severe headache; retro-orbital pain; muscle, joint, and bone pain; macular or maculopapular rash; and minor hemorrhagic manifestations, including petechiae, ecchymosis, purpura, epistaxis, bleeding gums, hematuria, or a positive tourniquet test result.[51] A recent systematic review and meta-analysis showed that allergic symptoms are one of the core symptoms that are highly associated with dengue severity.[52]
Mechanism of infection
- Dengue virus' (DENV) E envelope protein binds to a cellular receptor. The exact nature of the cellular receptor has not been fully elucidated.
- DENV undergoes endocytosis. Acidification of the endosome leads to a conformational change of E, exposing a fusion peptide sequence that facilitates fusion of the envelope with the endosomal membrane, releasing the virion capsid into the cytoplasm.
- Uncoating occurs in the cytoplasm
- Host translational machinery (ribosomes) translates the (+)ssRNA into a single polypeptide
- Cellular and viral proteinases cleave the polypeptide into 10 proteins (E, M, C and 7 nonstructural/enzymatic proteins) while embedded on the ER membrane.
- As soon as functional RNA-dependent RNA polymerase is synthesized, RNA replication can commence. Synthesis is asymmetrical, making 10 times more of the positive-sense strand than the negative.
- Assembly occurs on intracellular membranes, which bud into the ER (forming the envelope from the ER membrane). Subsequent budding from the ER through the Golgi and into vesicles allows maturation via posttranslational modifications, e.g. glycosylation and pH transformational rearrangements
- Egress occurs via exocytosis[53]
Severe disease
Some people develop more severe forms of dengue, such as dengue hemorrhagic fever. Different strains of viruses interacting with people with different immune backgrounds lead to a complex interaction. Among the possible causes are cross-serotypic immune response, through a mechanism known as antibody-dependent enhancement, which happens when a person who has been previously infected with dengue gets infected for the second, third, or fourth time. The previous antibodies to the old strain of dengue virus now interfere with the immune response to the current strain, leading paradoxically to more virus entry and uptake.[54]
Immune system interaction
In recent years, many studies have shown that flaviviruses, especially dengue virus, has the ability to inhibit the innate immune response during the infection.[55][56] Indeed, dengue virus has many nonstructural proteins that allow the inhibition of various mediators of the innate immune system response. These proteins act on two levels :
Inhibition of interferon signaling by blocking signal transducer
NS4B is a small, hydrophobic protein located in association with the endoplasmic reticulum. It may block the phosphorylation of STAT 1 after induction by interferons type I alpha & beta. In fact, as the activity of Tyk2 kinase decreases in association with dengue virus, so too does STAT 1 phosphorylation.[57] Furthermore, the innate immune system's response to the virus is further damped as expression of interferon-stimulating gene(s) (ISG) is restricted by the aforementioned 'NS4B' protein. NS2A and NS4A cofactor may also take part in the STAT 1 inhibition.[58]
NS5 - the presence of this 105-kDa protein results in inactivation of STAT2 (via the signal transduction of the response to interferon) when it is expressed alone.[59] When NS5 is cleaved with NS4B by a protease (NS2B3), it can degrade STAT2. In fact, after the cleavage of NS5 by the protease, an E3 ligase association with STAT2 occurs, and the E3 ligase targets STAT2 for the degradation.[60][61]
Inhibition of the type I interferon response
NS2B3-b protease complex is a proteolytic core consisting of the last 40 amino acids of NS2B and the first 180 amino acids of NS3. Cleavage of the NS2B3 precursor activates the protease complex.[62]
This protease complex allows the inhibition of the production of type I interferon by reducing the activity of IFN-beta promoter; NS2B3 protease complex is involved in inhibiting the phosphorylation of IRF3.[63] The NS2B3 protease complex inhibits (by cleaving) protein MITA which allows the IRF3 activation.[64]
Mosquito D7 saliva protein
Dengue virus is transmitted by the mosquito species Aedes aegypti, which produces saliva that contains over 100 unique proteins, including the protein family D7.[65] Scientists used to believe that A. aegypti saliva, when being transmitted, actually enhanced dengue virus in the body. The mosquito's saliva was thought to make the virus spread faster due to the weakened immune response of its host. However, a current study has found that the protein D7 hinders the virus transmission into the host cells.[65]
The immune responses of antibodies that are trying to fight off the foreign virus actually increase transmission and make the infection worse. Levels of protein D7 are more prevalent in salivary glands of dengue-infected mosquitoes compared to those uninfected ones.[65] D7 is found in mosquito saliva and was thought to assist the process of blood feeding. Despite the prior assumptions, D7 can modulate the host cell and act against the virus to prevent viral infection.[65] Unfortunately, D7 proteins provoke immune responses, which raise anti-D7 antibody levels. These antibodies inhibit the function of D7 proteins, which enhance transmission of dengue virus. Although immune responses against D7 proteins might impair their antiviral activity, a study showed that non-DENV subjects have slightly higher anti-D7 IgG levels than infected ones, although it was not statistically significant.[66] Thus, more studies over D7 protein family are needed do elucidate its role on DENV infection and its applicability in medicine.
Vaccine research
Only one vaccine for dengue is currently approved in 11 countries (Mexico, the Philippines, Indonesia, Brazil, El Salvador, Costa Rica, Paraguay, Guatemala, Peru, Thailand, and Singapore).[67][68][69] Several vaccines are under development by private and public researchers.[70] Developing a vaccine against the disease is challenging. With four different serotypes of the virus that can cause the disease, the vaccine must immunize against all four types to be effective.[3] Vaccination against only one serotype could possibly lead to severe dengue hemorrhagic shock when infected with another serotype due to antibody-dependent enhancement. When infected with dengue virus, the immune system produces cross-reactive antibodies that provide immunity to that particular serotype. However, these antibodies are incapable of neutralizing other serotypes upon reinfection and actually increase viral replication. When macrophages consume the 'neutralized' virus, the virus is able to replicate within the macrophage, causing disease. These cross-reactive, ineffective antibodies ease access of virus into macrophages, which induces more severe disease (dengue hemorrhagic fever, dengue shock syndrome). A common problem faced in dengue-endemic regions is when mothers become infected with dengue; after giving birth, offspring carry the immunity from their mother and are susceptible to hemorrhagic fever if infected with any of the other three serotypes.[71] One vaccine was in phase III trials in 2012 and planning for vaccine usage and effectiveness surveillance had started.[72]
In 2009, Sanofi-Pasteur started building a new facility in Neuville-sur-Saône' (fr), a suburb of Lyon (France). This unit produces four-serotype vaccine for phase III trials. In September 2014, the Sanofi-Pasteur CEO gave early results of the phase III trial efficacy study in Latin America. The efficacy per serotype (ST) varied widely, 42.3% for ST2, 50.3% for ST1, 74.0% for ST3, and 77.7% for ST4. The full analysis of data from the phase III Latin American-Caribbean study will be reviewed by external experts before being published in a peer-reviewed scientific journal. Primary results has to be presented at the American Society of Tropical Medicine and Hygiene Annual Meeting, held November 2–6, 2014, in New Orleans.[73]
In September 2012, one of the vaccines was reported to not have done well in clinical trials.[3]
In late 2015 and early 2016, the first dengue vaccine, Dengvaxia (CYD-TDV) by Sanofi-Pasteur, was registered in several countries for use in individuals 9–45 years of age living in endemic areas.
On May 1, 2019, the U.S. Food and Drug Administration announced the approval of Dengvaxia, the first vaccine for the prevention of dengue disease caused by all dengue virus serotypes in people ages 9 through 16 who have laboratory-confirmed previous dengue infection and who live in endemic areas. Dengue is endemic in the U.S. territories of American Samoa, Guam, Puerto Rico, and the U.S. Virgin Islands.[74]
Drug research
There are no approved direct antiviral treatments for Dengue fever. Most antiviral drug research for Dengue infections has focussed on inhibition of the NS2B/NS3 protease or NS5 proteins. Reported protease inhibitor approaches have focussed mainly on targeted covalent inhibitors.[75][76] One drug, Balapiravir, a repurposed hepatitis C NS5 polymerase inhibitor progressed to a Phase II clinical trial before being stopped due to lack of efficacy.[77][78]
References
- Rodenhuis-Zybert IA, Wilschut J, Smit JM (August 2010). "Dengue virus life cycle: viral and host factors modulating infectivity". Cellular and Molecular Life Sciences. 67 (16): 2773–86. doi:10.1007/s00018-010-0357-z. PMID 20372965. S2CID 4232236.
- WHO (2009). Dengue Guidelines for Diagnosis, Treatment, Prevention and Control (PDF). World Health Organization. ISBN 978-92-4-154787-1.
- Normile D (October 2013). "Tropical medicine. Surprising new dengue virus throws a spanner in disease control efforts". Science. 342 (6157): 415. Bibcode:2013Sci...342..415N. doi:10.1126/science.342.6157.415. PMID 24159024.
- Dwivedi, V. D., Tripathi, I. P., Tripathi, R. C., Bharadwaj, S., & Mishra, S. K. (2017). Genomics, proteomics and evolution of Dengue virus. Briefings in functional genomics.16(4): 217–227, https://doi.org/10.1093/bfgp/elw040
- Taylor-Robinson, Andrew W. (2016-03-03). "A Putative Fifth Serotype of Dengue - Potential Implications for Diagnosis, Therapy and Vaccine Design". International Journal of Clinical & Medical Microbiology. 2016. doi:10.15344/2456-4028/2016/101. ISSN 2456-4028.
- "A new understanding of Dengue virus". ScienceDaily. September 18, 2015. Retrieved May 7, 2016.
- "A Second Time Dengue Infection Can be Life-Threatening Says Research". NDTV. United Kingdom. September 18, 2015. Retrieved 2016-05-07.
- Vogels CB, Rückert C, Cavany SM, Perkins TA, Ebel GD, Grubaugh ND (January 2019). "Arbovirus coinfection and co-transmission: A neglected public health concern?". PLOS Biology. 17 (1): e3000130. doi:10.1371/journal.pbio.3000130. PMC 6358106. PMID 30668574.
- Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB, Jaenisch T, Wint GR, Simmons CP, Scott TW, Farrar JJ, Hay SI (April 2013). "The global distribution and burden of dengue". Nature. 496 (7446): 504–7. Bibcode:2013Natur.496..504B. doi:10.1038/nature12060. PMC 3651993. PMID 23563266.
- Mustafa, M. S.; Rasotgi, V.; Jain, S.; Gupta, V. (2015). "Discovery of fifth serotype of dengue virus (DENV-5): A new public health dilemma in dengue control". Medical Journal, Armed Forces India. 71 (1): 67–70. doi:10.1016/j.mjafi.2014.09.011. ISSN 0377-1237. PMC 4297835. PMID 25609867.
- Costa RL, Voloch CM, Schrago CG (2012) Comparative evolutionary epidemiology of dengue virus serotypes. Infect Genet Evol 12(2):309-314
- "Dengue virus". Pathogen Information (PathInfo). Virginia Bioinformatics Institute, Virginia Tech. Archived from the original on 2010-08-30.
- Holmes EC, Twiddy SS (May 2003). "The origin, emergence and evolutionary genetics of dengue virus". Infection, Genetics and Evolution. 3 (1): 19–28. doi:10.1016/s1567-1348(03)00004-2. PMID 12797969.
- Halstead SB (January 1988). "Pathogenesis of dengue: challenges to molecular biology". Science. 239 (4839): 476–81. Bibcode:1988Sci...239..476H. doi:10.1126/science.3277268. PMID 3277268.
- Haddow AD, Guzman H, Popov VL, Wood TG, Widen SG, Haddow AD, Tesh RB, Weaver SC (June 2013). "First isolation of Aedes flavivirus in the Western Hemisphere and evidence of vertical transmission in the mosquito Aedes (Stegomyia) albopictus (Diptera: Culicidae)". Virology. 440 (2): 134–9. doi:10.1016/j.virol.2012.12.008. PMID 23582303.
- Thongyuan S, Kittayapong P (30 August 2017). "First evidence of dengue infection in domestic dogs living in different ecological settings in Thailand". PLOS ONE. 12 (8): e0180013. Bibcode:2017PLoSO..1280013T. doi:10.1371/journal.pone.0180013. PMC 5576688. PMID 28854207.
- Ghosh Roy S, Sadigh B, Datan E, Lockshin RA, Zakeri Z (May 2014). "Regulation of cell survival and death during Flavivirus infections". World Journal of Biological Chemistry. 5 (2): 93–105. doi:10.4331/wjbc.v5.i2.93 (inactive 31 July 2022). PMC 4050121. PMID 24921001.
{{cite journal}}: CS1 maint: DOI inactive as of July 2022 (link) - Datan E, Roy SG, Germain G, Zali N, McLean JE, Golshan G, Harbajan S, Lockshin RA, Zakeri Z (March 2016). "Dengue-induced autophagy, virus replication and protection from cell death require ER stress (PERK) pathway activation". Cell Death & Disease. 7 (e2127): e2127. doi:10.1038/cddis.2015.409. PMC 4823927. PMID 26938301.
- McLean JE, Wudzinska A, Datan E, Quaglino D, Zakeri Z (June 2011). "Flavivirus NS4A-induced autophagy protects cells against death and enhances virus replication". The Journal of Biological Chemistry. 286 (25): 22147–59. doi:10.1074/jbc.M110.192500. PMC 3121359. PMID 21511946.
- Zakeri Z, Ghosh-Roy S, Datan E, McLean JE, Zandi K, Germain G (2015). "Cell death and virus infection – a short review". 20 Years of Cell Death. 1 (1): 292–325. doi:10.13140/RG.2.2.35359.97447.
- Neufeldt CJ, Cortese M, Acosta EG, Bartenschlager R (February 2018). "Rewiring cellular networks by members of the Flaviviridae family". Nature Reviews. Microbiology. 16 (3): 125–142. doi:10.1038/nrmicro.2017.170. PMC 7097628. PMID 29430005.
- Hanley KA, Weaver SC, eds. (2010). Frontiers in Dengue Virus Research. Caister Academic. p. 13. ISBN 978-1-904455-50-9. Retrieved 22 September 2020.
- Modis Y, Ogata S, Clements D, Harrison SC (June 2003). "A ligand-binding pocket in the dengue virus envelope glycoprotein". Proceedings of the National Academy of Sciences of the United States of America. 100 (12): 6986–91. Bibcode:2003PNAS..100.6986M. doi:10.1073/pnas.0832193100. PMC 165817. PMID 12759475.
- Perera R, Kuhn RJ (August 2008). "Structural proteomics of dengue virus". Current Opinion in Microbiology. 11 (4): 369–77. doi:10.1016/j.mib.2008.06.004. PMC 2581888. PMID 18644250.
- Tassaneetrithep B, Burgess TH, Granelli-Piperno A, Trumpfheller C, Finke J, Sun W, Eller MA, Pattanapanyasat K, Sarasombath S, Birx DL, Steinman RM, Schlesinger S, Marovich MA (April 2003). "DC-SIGN (CD209) mediates dengue virus infection of human dendritic cells". The Journal of Experimental Medicine. 197 (7): 823–9. doi:10.1084/jem.20021840. PMC 2193896. PMID 12682107.
- Krishnan MN, Sukumaran B, Pal U, Agaisse H, Murray JL, Hodge TW, Fikrig E (May 2007). "Rab 5 is required for the cellular entry of dengue and West Nile viruses". Journal of Virology. 81 (9): 4881–5. doi:10.1128/JVI.02210-06. PMC 1900185. PMID 17301152.
- Jindadamrongwech S, Thepparit C, Smith DR (May 2004). "Identification of GRP 78 (BiP) as a liver cell expressed receptor element for dengue virus serotype 2". Archives of Virology. 149 (5): 915–27. doi:10.1007/s00705-003-0263-x. PMID 15098107. S2CID 12062084.
- Miller JL, de Wet BJ, deWet BJ, Martinez-Pomares L, Radcliffe CM, Dwek RA, Rudd PM, Gordon S (February 2008). "The mannose receptor mediates dengue virus infection of macrophages". PLOS Pathogens. 4 (2): e17. doi:10.1371/journal.ppat.0040017. PMC 2233670. PMID 18266465.
- Zidane N, Ould-Abeih MB, Petit-Topin I, Bedouelle H (December 2012). "The folded and disordered domains of human ribosomal protein SA have both idiosyncratic and shared functions as membrane receptors". Bioscience Reports. 33 (1): 113–24. doi:10.1042/BSR20120103. PMC 4098866. PMID 23137297.
- Hussain, Mazhar; Idrees, Muhammad; Afzal, Samia (2015-07-22). "Development of Global Consensus of Dengue Virus Envelope Glycoprotein for Epitopes Based Vaccine Design". Current Computer-Aided Drug Design. 11 (1): 84–97. doi:10.2174/1573409911666150529130134. ISSN 1573-4099. PMID 26022067.
- Zidane N, Dussart P, Bremand L, Bedouelle H (July 2013). "Cross-reactivities between human IgMs and the four serotypes of dengue virus as probed with artificial homodimers of domain-III from the envelope proteins". BMC Infectious Diseases. 13: 302. doi:10.1186/1471-2334-13-302. PMC 3701519. PMID 23815496.
- Zidane N, Dussart P, Bremand L, Villani ME, Bedouelle H (June 2013). "Thermodynamic stability of domain III from the envelope protein of flaviviruses and its improvement by molecular design". Protein Engineering, Design & Selection. 26 (6): 389–99. doi:10.1093/protein/gzt010. PMID 23479674.
- Brandler S, Ruffie C, Najburg V, Frenkiel MP, Bedouelle H, Desprès P, Tangy F (September 2010). "Pediatric measles vaccine expressing a dengue tetravalent antigen elicits neutralizing antibodies against all four dengue viruses". Vaccine. 28 (41): 6730–9. doi:10.1016/j.vaccine.2010.07.073. PMID 20688034.
- Miller S, Kastner S, Krijnse-Locker J, Bühler S, Bartenschlager R (March 2007). "The non-structural protein 4A of dengue virus is an integral membrane protein inducing membrane alterations in a 2K-regulated manner". The Journal of Biological Chemistry. 282 (12): 8873–82. doi:10.1074/jbc.M609919200. PMID 17276984. S2CID 19927382.
- Lee CM, Xie X, Zou J, Li SH, Lee MY, Dong H, Qin CF, Kang C, Shi PY (June 2015). "Determinants of Dengue Virus NS4A Protein Oligomerization". Journal of Virology. 89 (12): 6171–83. doi:10.1128/JVI.00546-15. PMC 4474302. PMID 25833044.
- Zou J, Xie X, Wang QY, Dong H, Lee MY, Kang C, Yuan Z, Shi PY (April 2015). "Characterization of dengue virus NS4A and NS4B protein interaction". Journal of Virology. 89 (7): 3455–70. doi:10.1128/JVI.03453-14. PMC 4403404. PMID 25568208.
- Austin SK, Dowd KA, Shrestha B, Nelson CA, Edeling MA, Johnson S, Pierson TC, Diamond MS, Fremont DH (2012). "Structural basis of differential neutralization of DENV-1 genotypes by an antibody that recognizes a cryptic epitope". PLOS Pathogens. 8 (10): e1002930. doi:10.1371/journal.ppat.1002930. PMC 3464233. PMID 23055922.
- Lok SM, Kostyuchenko V, Nybakken GE, Holdaway HA, Battisti AJ, Sukupolvi-Petty S, Sedlak D, Fremont DH, Chipman PR, Roehrig JT, Diamond MS, Kuhn RJ, Rossmann MG (March 2008). "Binding of a neutralizing antibody to dengue virus alters the arrangement of surface glycoproteins". Nature Structural & Molecular Biology. 15 (3): 312–7. doi:10.1038/nsmb.1382. PMID 18264114. S2CID 64886.
- Midgley CM, Flanagan A, Tran HB, Dejnirattisai W, Chawansuntati K, Jumnainsong A, Wongwiwat W, Duangchinda T, Mongkolsapaya J, Grimes JM, Screaton GR (May 2012). "Structural analysis of a dengue cross-reactive antibody complexed with envelope domain III reveals the molecular basis of cross-reactivity". Journal of Immunology. 188 (10): 4971–9. doi:10.4049/jimmunol.1200227. PMC 3364712. PMID 22491255.
- Lisova O, Hardy F, Petit V, Bedouelle H (September 2007). "Mapping to completeness and transplantation of a group-specific, discontinuous, neutralizing epitope in the envelope protein of dengue virus". The Journal of General Virology. 88 (Pt 9): 2387–97. doi:10.1099/vir.0.83028-0. PMID 17698647.
- Cockburn JJ, Navarro Sanchez ME, Fretes N, Urvoas A, Staropoli I, Kikuti CM, Coffey LL, Arenzana Seisdedos F, Bedouelle H, Rey FA (February 2012). "Mechanism of dengue virus broad cross-neutralization by a monoclonal antibody". Structure. 20 (2): 303–14. doi:10.1016/j.str.2012.01.001. PMID 22285214.
- Lisova O, Belkadi L, Bedouelle H (April 2014). "Direct and indirect interactions in the recognition between a cross-neutralizing antibody and the four serotypes of dengue virus". Journal of Molecular Recognition. 27 (4): 205–14. doi:10.1002/jmr.2352. PMID 24591178. S2CID 5416842.
- Cockburn JJ, Navarro Sanchez ME, Goncalvez AP, Zaitseva E, Stura EA, Kikuti CM, Duquerroy S, Dussart P, Chernomordik LV, Lai CJ, Rey FA (February 2012). "Structural insights into the neutralization mechanism of a higher primate antibody against dengue virus". The EMBO Journal. 31 (3): 767–79. doi:10.1038/emboj.2011.439. PMC 3273384. PMID 22139356.
- Rouvinski A, Guardado-Calvo P, Barba-Spaeth G, Duquerroy S, Vaney MC, Kikuti CM, Navarro Sanchez ME, Dejnirattisai W, Wongwiwat W, Haouz A, Girard-Blanc C, Petres S, Shepard WE, Desprès P, Arenzana-Seisdedos F, Dussart P, Mongkolsapaya J, Screaton GR, Rey FA (April 2015). "Recognition determinants of broadly neutralizing human antibodies against dengue viruses" (PDF). Nature. 520 (7545): 109–13. Bibcode:2015Natur.520..109R. doi:10.1038/nature14130. hdl:10044/1/34173. PMID 25581790. S2CID 4394235.
- Wiatrek, Melissa. Dengue Fever. http://www.austincc.edu/microbio/2993p/df.htm Archived 2018-07-26 at the Wayback Machine
- World Health Organization. Dengue and severe dengue. April 2017. http://www.who.int/mediacentre/factsheets/fs117/en/
- DISTRICT OF COLUMBIA DEPARTMENT OF HEALTH. Dengue Fever Factsheet. https://doh.dc.gov/sites/default/files/dc/sites/doh/publication/attachments/Dengue_Fever.pdf
- Better Health. Dengue virus disease. https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/Dengue-virus-disease?viewAsPdf=true
- Centers for Disease Control and Prevention. Epidemiology. https://www.cdc.gov/dengue/epidemiology/
- Centers for Disease Control and Prevention. Dengue. https://wwwnc.cdc.gov/travel/yellowbook/2016/infectious-diseases-related-to-travel/dengue
- Kien ND, El-Qushayri AE, Ahmed AM, Safi A, Mageed SA, Mehyar SM, et al. (February 2020). "Association of Allergic Symptoms with Dengue Infection and Severity: A Systematic Review and Meta-analysis". Virologica Sinica. 35 (1): 83–92. doi:10.1007/s12250-019-00165-6. PMC 7035405. PMID 31637633.
- Acheson, Nicholas H. (2011). Fundamentals of Molecular Virology, 2nd ed. Wiley.
- Dejnirattisai W, Jumnainsong A, Onsirisakul N, Fitton P, Vasanawathana S, Limpitikul W, Puttikhunt C, Edwards C, Duangchinda T, Supasa S, Chawansuntati K, Malasit P, Mongkolsapaya J, Screaton G (May 2010). "Cross-reacting antibodies enhance dengue virus infection in humans". Science. 328 (5979): 745–8. Bibcode:2010Sci...328..745D. doi:10.1126/science.1185181. PMC 3837288. PMID 20448183.
- Diamond MS (September 2009). "Mechanisms of evasion of the type I interferon antiviral response by flaviviruses". Journal of Interferon & Cytokine Research. 29 (9): 521–30. doi:10.1089/jir.2009.0069. PMID 19694536.
- Jones M, Davidson A, Hibbert L, Gruenwald P, Schlaak J, Ball S, Foster GR, Jacobs M (May 2005). "Dengue virus inhibits alpha interferon signaling by reducing STAT2 expression". Journal of Virology. 79 (9): 5414–20. doi:10.1128/JVI.79.9.5414-5420.2005. PMC 1082737. PMID 15827155.
- Ho LJ, Hung LF, Weng CY, Wu WL, Chou P, Lin YL, Chang DM, Tai TY, Lai JH (June 2005). "Dengue virus type 2 antagonizes IFN-alpha but not IFN-gamma antiviral effect via down-regulating Tyk2-STAT signaling in the human dendritic cell". Journal of Immunology. 174 (12): 8163–72. doi:10.4049/jimmunol.174.12.8163. PMID 15944325. S2CID 83577905.
- Muñoz-Jordan JL, Sánchez-Burgos GG, Laurent-Rolle M, García-Sastre A (November 2003). "Inhibition of interferon signaling by dengue virus". Proceedings of the National Academy of Sciences of the United States of America. 100 (24): 14333–8. Bibcode:2003PNAS..10014333M. doi:10.1073/pnas.2335168100. PMC 283592. PMID 14612562.
- Ashour J, Laurent-Rolle M, Shi PY, García-Sastre A (June 2009). "NS5 of dengue virus mediates STAT2 binding and degradation". Journal of Virology. 83 (11): 5408–18. doi:10.1128/JVI.02188-08. PMC 2681973. PMID 19279106.
- Mazzon M, Jones M, Davidson A, Chain B, Jacobs M (October 2009). "Dengue virus NS5 inhibits interferon-alpha signaling by blocking signal transducer and activator of transcription 2 phosphorylation". The Journal of Infectious Diseases. 200 (8): 1261–70. doi:10.1086/605847. PMID 19754307.
- Morrison J, Aguirre S, Fernandez-Sesma A (March 2012). "Innate immunity evasion by Dengue virus". Viruses. 4 (3): 397–413. doi:10.3390/v4030397. PMC 3347034. PMID 22590678.
- Yusof R, Clum S, Wetzel M, Murthy HM, Padmanabhan R (April 2000). "Purified NS2B/NS3 serine protease of dengue virus type 2 exhibits cofactor NS2B dependence for cleavage of substrates with dibasic amino acids in vitro". The Journal of Biological Chemistry. 275 (14): 9963–9. doi:10.1074/jbc.275.14.9963. PMID 10744671. S2CID 30345627.
- Rodriguez-Madoz JR, Belicha-Villanueva A, Bernal-Rubio D, Ashour J, Ayllon J, Fernandez-Sesma A (October 2010). "Inhibition of the type I interferon response in human dendritic cells by dengue virus infection requires a catalytically active NS2B3 complex". Journal of Virology. 84 (19): 9760–74. doi:10.1128/JVI.01051-10. PMC 2937777. PMID 20660196.
- Yu CY, Chang TH, Liang JJ, Chiang RL, Lee YL, Liao CL, Lin YL (June 2012). Diamond MS (ed.). "Dengue virus targets the adaptor protein MITA to subvert host innate immunity". PLOS Pathogens. 8 (6): e1002780. doi:10.1371/journal.ppat.1002780. PMC 3386177. PMID 22761576.
- Conway MJ, Londono-Renteria B, Troupin A, Watson AM, Klimstra WB, Fikrig E, Colpitts TM (September 2016). "Aedes aegypti D7 Saliva Protein Inhibits Dengue Virus Infection". PLOS Neglected Tropical Diseases. 10 (9): e0004941. doi:10.1371/journal.pntd.0004941. PMC 5025043. PMID 27632170.
- Londono-Renteria BL, Shakeri H, Rozo-Lopez P, Conway MJ, Duggan N, Jaberi-Douraki M, Colpitts TM (2018). "Aedes aegypti D7 Salivary Proteins in Colombia". Frontiers in Public Health. 6: 111. doi:10.3389/fpubh.2018.00111. PMC 5968123. PMID 29868532.
- "Sanofi's dengue vaccine approved in 11 countries". Reuters. 2016. Retrieved 13 August 2017.
- East, Susie (6 April 2016). "World's first dengue fever vaccine launched in the Philippines". CNN. Retrieved 17 October 2016.
- "Dengue Fever Vaccine Available in Indonesia". October 17, 2016.
- Vaccine Development Archived 2019-08-21 at the Wayback Machine, Dengue Vaccine Initiative, November 2012, accessed November 5, 2013
- Schmaljohn AL, McClain D (1996). "Ch. 54: Alphaviruses (Togaviridae) and Flaviviruses (Flaviviridae)". In Baron S (ed.). Medical Microbiology (4th ed.). University of Texas Medical Branch at Galveston. ISBN 978-0-9631172-1-2. NBK7633.
- Torresi J, Tapia-Conyer R, Margolis H (2013). "Preparing for dengue vaccine introduction: recommendations from the 1st dengue v2V International Meeting". PLOS Neglected Tropical Diseases. 7 (9): e2261. doi:10.1371/journal.pntd.0002261. PMC 3784468. PMID 24086776.
- "Sanofi Pasteur Dengue Vaccine Aces Second Phase III Trial". GEN News Highlights. Genetic Engineering & Biotechnology News. 3 September 2014.
- First FDA-approved vaccine for the prevention of dengue disease in endemic regions https://www.fda.gov/news-events/press-announcements/first-fda-approved-vaccine-prevention-dengue-disease-endemic-regions
- Leung, D.; Schroder, K.; White, H.; Fang, N. X.; Stoermer, M. J.; Abbenante, G.; Martin, J. L.; Young, P. R.; Fairlie, D. P. (2001-12-07). "Activity of recombinant dengue 2 virus NS3 protease in the presence of a truncated NS2B co-factor, small peptide substrates, and inhibitors". The Journal of Biological Chemistry. 276 (49): 45762–45771. doi:10.1074/jbc.M107360200. ISSN 0021-9258. PMID 11581268. S2CID 26124089.
- Dražić, Tonko; Kopf, Sara; Corridan, James; Leuthold, Mila M.; Bertoša, Branimir; Klein, Christian D. (2020-01-09). "Peptide-β-lactam Inhibitors of Dengue and West Nile Virus NS2B-NS3 Protease Display Two Distinct Binding Modes". Journal of Medicinal Chemistry. 63 (1): 140–156. doi:10.1021/acs.jmedchem.9b00759. ISSN 0022-2623. PMID 31804823. S2CID 208768169.
- Nguyen NM, Tran CN, Phung LK, Duong KT, Huynh HA, Farrar J, et al. (May 2013). "A randomized, double-blind placebo controlled trial of balapiravir, a polymerase inhibitor, in adult dengue patients". The Journal of Infectious Diseases. 207 (9): 1442–50. doi:10.1093/infdis/jis470. PMC 3610419. PMID 22807519.
- Chen YL, Abdul Ghafar N, Karuna R, Fu Y, Lim SP, Schul W, et al. (February 2014). "Activation of peripheral blood mononuclear cells by dengue virus infection depotentiates balapiravir". Journal of Virology. 88 (3): 1740–7. doi:10.1128/JVI.02841-13. PMC 3911617. PMID 24257621.
External links
- 3D electron microscopy structures of dengue virus from the EM Data Bank(EMDB)
- "Brazil releases 'good' mosquitoes to fight dengue fever". BBC News Latin America & Caribbean. 24 September 2014.