Murray's law
In biophysical fluid dynamics, Murray's law is a potential relationship between radii at junctions in a network of fluid-carrying tubular pipes. Its simplest version proposes that whenever a branch of radius splits into two branches of radii and , then all three radii should obey the equation




If network flow is smooth and leak-free, then systems that obey Murray's law minimize the resistance to flow through the network. For turbulent networks, the law takes the same form but with a different characteristic exponent α.
Murray's law is observed in the vascular and respiratory systems of animals, xylem in plants, and the respiratory system of insects. In principle, Murray's law also applies to biomimetic engineering, but human designs rarely exploit the law.
Murray's law is named after Cecil D. Murray, a physiologist at Bryn Mawr College, who first argued that efficient transport might determine the structure of the human vascular system.
Assumptions
Murray's law assumes material is passively transported by the flow of fluid in a network of tubular pipes,[1] and that said network requires energy both to maintain flow and structural integrity.[2] Variation in the fluid viscosity across scales will affect the Murray's law exponent, but is usually too small to matter.[3]
At least two different conditions are known in which the cube exponent is optimal.
In the first, organisms have free (variable) circulatory volume. Also, maintenance energy is not proportional to the pipe material, but instead the quantity of working fluid. The latter assumption is justified in metabolically active biological fluids, such as blood.[4] It is also justified for metabolically inactive fluids, such as air, as long as the energetic "cost" of the infrastructure scales with the cross-sectional area of each tube; such is the case for all known biological tubules.[5]
In the second, organisms have fixed circulatory volume and pressure, but wish to minimize the resistance to flow through the system. Equivalently, maintenance is negligible and organisms with to maximize the volumetric flow rate.[6]
Although most derivations of Murray's law assume a steady state flow field, the same results apply for flow in tubes that have a width moderate-to-small relative to the flow wavelength.[7]
Derivation in laminar, mass-conservative networks with non-negligible upkeep
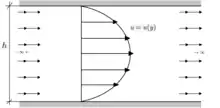
Murray's original derivation uses the first set of assumptions described above. She begins with the Hagen–Poiseuille equation, which states that for fluid of dynamic viscosity μ, flowing laminarly through a cylindrical pipe of radius r and length l, the volumetric flow rate Q associated with a pressure drop Δp is

and the corresponding power consumed is[1]

Said pipe contains volume πlr2. If the maintenance power density is λ, then the total power consumed (from both flow and upkeep) is

Minimizing this quantity depends on precisely which variables the organism is free to manipulate, but the minimum invariably occurs when the two terms are proportional to each other.[8] In that minimal case, the proportionality determines a relationship between Q and r. Canceling common factors and taking a square root,
-
(1)

That is, when using as little energy as possible, the mass flowing through the pipe must be proportional to the cube of the pipe's radius. Since flow is leakless, the total flow rate into a junction must be the total flow rate out:

Exponents for other networks
If the network does not rely on transported material getting "swept up in the flow", but instead expects it to passively diffuse, then resistance to transport is minimized when α=2: that is,

The same law would apply to a direct-current electrical grid composed of wires of only one material, but varying diameter.[10] For turbulent flow, transport resistance is minimized when α=7/3; that is:[11][12]

In general, networks intermediate between diffusion and laminar flow are expected to have characteristic exponents between 2 and 3, at least approximately.[13][14]
Experimental tests
Murray's law has been verified in chicks; dog intestines and lungs; cat mesentery; and human intestines and lung capillaries.[15][16] Mice genetically engineered to lack the blood-vessel-wall protein elastin have smaller and thinner blood vessels, but still obey Murray's law.[17]
In humans, large vessels, such as the aorta or trachea, do not appear to obey Murray's law, instead obeying a Murray's law with exponent close to 2.[16] But flow in those vessels is also partially turbulent, and so should exhibit an exponent nearer to 7/3 than to 3.[18]
_connecting_to_many_children_pores_(radius_ri)._.jpg.webp)
Insects do not have a fully-fledged circulatory system, instead relying on passive diffusion through the haemocoel. For those networks, Murray's law predicts constant cross-sectional area, which is observed.[10]
The same arguments that imply Murray's law also imply that the distribution of tubules should exhibit a specific power law scaling with size. Plant xylem is known to exhibit that scaling except in scenarios where the passages double as structural supports.[19][20]
History
The first phenomenon now recognized as Murray's law is Young's rule for circulatory systems, which states that two identical subcapillaries should combine to form a capillary with radius about 1.26≈3√2 times larger, and dates to the early 19th century.[21] Bryn Mawr physiologist Cecil D. Murray published the law's modern, general formulation in 1926,[22][21] but it languished in a disciplinary no-man's-land for the next fifty years: too trivial for physicists and too complicated for biologists.[21] Interest in the law revived in the 1970s.[21]
Applications
In circulatory system governed by Murray's law with α=3, shear stress on vessel walls is roughly constant. Consequently, variations in shear stress are a sign of deviation from Murray's law; Rodbard and Zamir suggest that such variations stimulate homeostatic growth or contraction.[23]
Murray's law rarely applies to engineered materials, because man-made transport routes attempt to reduce flow resistance by minimizing branching and maximizing diameter.[24]
Murray materials
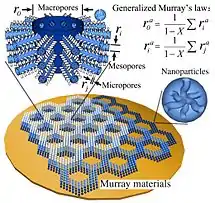
Materials that obey Murray's law at the microscale, known as Murray materials, are expected to have favorable flow characteristics, but their construction is difficult, because it requires tight control over pore size typically over a wide range of scales.[14][25]
Lim et al propose designing microfluidic "labs on a chip" in accord with Murray's law to minimize flow resistance during analysis. Conventional lithography does not support such construction, because it cannot produce channels of varying depth.[26]
Seeking long-lived lithium battery electrodes, Zheng et al constructed Murray materials out of layers of sintered zinc oxide nanoparticles. The evaporation rate of the dissolved zinc oxide solvent controlled the size of the pores in each layer; the network was then just layers of ZnO with different pore sizes placed atop each other.[14]
Because power plant working fluids typically funnel into many small tubules for efficient heat transfer, Murray's law may be appropriate for nuclear reactor design.[27]
References
Footnotes
- Murray 1926a, p. 210: "Poisseuille's equation for the law of flow in cylindrical tubes may be taken as a very fair approximation giving the relationships required."
- Murray 1926b, p. 299: "The cost per second, E, of operating a section of blood vessel was taken to be the sum of two work terms: (1) the work done per second against friction by the flow of blood, given in ergs/sec. by the product pf, where p is the fall in pressure in dynes/cm., and f is the rate of blood flow in cc./sec., and (2) the work done per second in maintaining the mere volume of blood contained in the vessel, given also in ergs/sec. by the term bvol, where vol is the volume in cc., and b is the cost in ergs/sec. per cc. of blood."
- Sherman 1981, p. 447.
- Murray 1926a, p. 210: "Let b, then, be the cost of blood in ergs per second per cubic centimeter of whole blood of average composition (and let B be the cost in Calories per day per cc. of blood). There is, as far as I can see, nothing arbitrary about this step: it is certain that the maintenance of blood requires fuel. (The cost of blood may, however, be a complex account distributed among such factors as the small metabolism of blood itself, the cost of upkeep of all the constituents, perhaps especially of hemoglobin, the cost of the containing vessels, and the burden placed upon the body in general by the mere weight of blood.)
- Sherman 1981, p. 437: "If the volume of vessel tissue is proportional to the square of the internal radius (as when wall thickness is proportional to r), then Murray's derivation can hold for a biological vasculature even when the flowing fluid itself is inert and nonliving, as in the airways of the lungs, where the vessels are filled with air (of no metabolic cost) rather than with blood."
- Sherman 1981, pp. 437–439.
- Painter, Edén & Bengtsson 2006.
- Murray 1926a, pp. 210, 212: equations (4) and (6). Murray 1926b, p. 304: "The primitive meaning of organization is contained in the simplest problem of maxima and minima: if xyz=E, the sum x+y+z, is a minimum when x=y=z=3√E.". Sherman 1981, p. 436: "Under these conditions, only the coefficient b changes by inclusion of the vessel wall; the overall derivation remains the same, and Murray's law still holds."
- Murray 1926a, p. 212: equation (7).
- Sherman 1981, p. 446.
- Uylings 1977, p. 514: equation (15) with the maximal value of j=5.
- Williams et al. 2008, p. 60: equation (5.2).
- Uylings 1977, p. 514: equation (15).
- Zheng et al. 2017.
- Williams et al. 2008, p. 56.
- Sherman 1981, pp. 440–445.
- Sather, Hageman & Wagenseil 2012.
- Uylings 1977, p. 509: "In the upper airways of the lung, turbulent flow exists during inspiration, which changes to laminar flow in the lower airways. The blood flow in the aorta of, for instance, normal rabbits and humans is turbulent under some circumstances."
- McCulloh, Sperry & Adler 2003.
- McCulloh & Sperry 2006.
- Sherman 1981, p. 432.
- Murray 1926a, 1926b.
- Sherman 1981, p. 449.
- Williams et al. 2008, p. 56: "This is not strictly accurate; Murray's law geometry was derived to obtain minimum system-level power demand and if lowest resistance is required then the largest vessel diameter should be selected at all points." The quote misidentifies the Murray's law optimization — Murray's law does provide lowest resistance, assuming system volume is fixed — but correctly identifies that, when system volume can grow freely, choosing maximal diameters will lower resistance.
- Williams et al. 2008.
- Lim et al. 2003.
- Acharya, Sarmistha (10 April 2017). "Leaf vein structure might help improve battery life". International Business Times. Yahoo! News. Archived from the original on 11 April 2017.
Further reading
- Murray, Cecil D. (1926a). "The Physiological Principle of Minimum Work: I. The Vascular System and the Cost of Blood Volume". Proceedings of the National Academy of Sciences of the United States of America. 12 (3): 207–214. Bibcode:1926PNAS...12..207M. doi:10.1073/pnas.12.3.207. PMC 1084489. PMID 16576980.
- Murray, Cecil D. (1926b). "The Physiological Principle of Minimum Work: II. Oxygen Exchange in Capillaries". Proceedings of the National Academy of Sciences of the United States of America. 12 (5): 299–304. Bibcode:1926PNAS...12..299M. doi:10.1073/pnas.12.5.299. PMC 1084544. PMID 16587082.
- Sherman, Thomas F. (1981). "On connecting large vessels to small: The meaning of Murray's law". The Journal of General Physiology. 78 (4): 431–53. doi:10.1085/jgp.78.4.431. PMC 2228620. PMID 7288393.
- Uylings, H. B. M. (1977). "Optimization of diameters and bifurcation angles in lung and vascular tree structures". Bulletin of Mathematical Biology. 39 (5): 509–520. doi:10.1007/BF02461198. ISSN 1522-9602. PMID 890164. S2CID 189884751.
- McCulloh, Katherine A.; Sperry, John S.; Adler, Frederick R. (2003). "Water transport in plants obeys Murray's law". Nature. 421 (6926): 939–942. Bibcode:2003Natur.421..939M. doi:10.1038/nature01444. PMID 12607000. S2CID 4395034.
- Lim, Daniel; Kamotani, Yoko; Cho, Brenda; Mazumder, Jyotirmoy; Takayama, Shuichi (2003). "Fabrication of microfluidic mixers and artificial vasculatures using a high-brightness diode-pumped Nd:YAG laser direct write method". Lab on a Chip. 3 (4): 318–323. doi:10.1039/B308452C. ISSN 1473-0189. PMID 15007466.
- McCulloh, Katherine A.; Sperry, John S. (2006). "Murray's Law and the Vascular Architecture of Plants". In Herrel, Anthony; Speck, Thomas; Rowe, Nicholas P. (eds.). Ecology and Biomechanics: A Mechanical Approach to the Ecology of Animals and Plants. pp. 85–100. doi:10.1201/9781420001594. ISBN 9780429122323.
- Painter, Page R; Edén, Patrik; Bengtsson, Hans-Uno (2006). "Pulsatile blood flow, shear force, energy dissipation and Murray's Law". Theoretical Biology and Medical Modelling. 3 (1): 31. doi:10.1186/1742-4682-3-31. ISSN 1742-4682. PMC 1590016. PMID 16923189.
- Williams, Hugo R.; Trask, Richard S.; Weaver, Paul M.; Bond, Ian P. (2008). "Minimum mass vascular networks in multifunctional materials". Journal of the Royal Society Interface. 5 (18): 55–65. doi:10.1098/rsif.2007.1022. PMC 2605499. PMID 17426011.
- Sather, Bradley A.; Hageman, Daniel; Wagenseil, Jessica E. (2012). "Murray's Law in Elastin Haploinsufficient (Eln+/-) and Wild-Type (WT) Mice". Journal of Biomechanical Engineering. 134 (12): 124504-1–124504-3. doi:10.1115/1.4023093. PMC 3644989. PMID 23363211.
- Zheng, Xianfeng; Shen, Guofang; Wang, Chao; Li, Yu; Dunphy, Darren; Hasan, Tawfique; Brinker, C. Jeffrey; Su, Bao-Lian (2017-04-06). "Bio-inspired Murray materials for mass transfer and activity". Nature Communications. 8: 14921. Bibcode:2017NatCo...814921Z. doi:10.1038/ncomms14921. ISSN 2041-1723. PMC 5384213. PMID 28382972.