Respiratory burst
Respiratory burst (or oxidative burst) is the rapid release of the reactive oxygen species (ROS), superoxide anion (O−
2) and hydrogen peroxide (H
2O
2), from different cell types.
This is usually utilised for mammalian immunological defence, but also plays a role in cell signalling. Respiratory burst is also implicated in the ovum of animals following fertilization. It may also occur in plant cells.
Immunity
Immune cells can be divided into myeloid cells and lymphoid cells. Myeloid cells, including macrophages and neutrophils, are especially implicated in the respiratory burst.[1] They are phagocytic, and the respiratory burst is vital for the subsequent degradation of internalised bacteria or other pathogens. This is an important aspect of the innate immunity.
Respiratory burst requires a 10 to 20 fold increase in oxygen consumption through NADPH oxidase (NOX2 in humans) activity. NADPH is the key substrate of NOX2, and bears reducing power. Glycogen breakdown is vital to produce NADPH. This occurs via the pentose phosphate pathway.
The NOX2 enzyme is bound in the phagolysosome membrane. Post bacterial phagocytosis, it is activated, producing superoxide via its redox centre, which transfers electrons from cytosolic NADPH to O2 in the phagosome.[2]
2O2 + NADPH —> 2O2•– + NADP+ + H+
The superoxide can then spontaneously or enzymatically react with other molecules to give rise to other ROS. The phagocytic membrane reseals to limit exposure of the extracellular environment to the generated reactive free radicals.
Pathways for reactive species generation
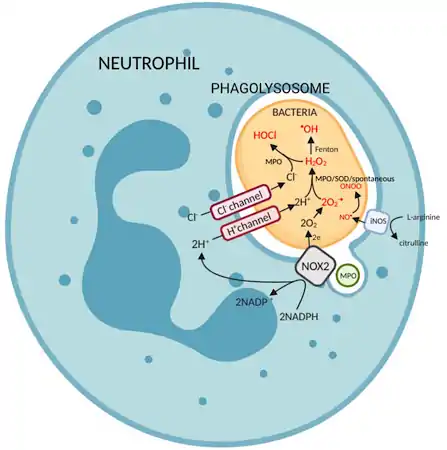
There are 3 main pathways for the generation of reactive oxygen species or reactive nitrogen species (RNS) in effector cells:[3]
- Superoxide dismutase (or alternatively, myeloperoxidase) generates hydrogen peroxide from superoxide. Hydroxyl radicals are then generated via the Haber-Weiss reaction or the Fenton reaction, of which are both catalyzed by Fe2+.
O2•–+ H2O2 —> •OH + OH– + O2 - In the presence of halide ions, prominently chloride ions, myeloperoxidase uses hydrogen peroxide to produce hypochlorous acid.
H2O2 + Cl− —> ClO− + H2O - Nitric oxide synthase (the inducible isoform, iNOS, in immunity) catalyses the production of nitric oxide from L-arginine. 2L-arginine + 3NADPH + 3 H+ + 4O2 —> 2citrulline + 2NO• + 4H2O + 3NADP+
Nitric oxide may react with superoxide anions to produce peroxynitrite anion.
Defense against pathogens
The exposure to these reactive species in the respiratory burst results in pathology. This is due to oxidative damage to the engulfed bacteria.
Notably, peroxynitrite is a very strong oxidising agent that can lead to lipid peroxidation, protein oxidation, protein nitration, which are responsible for its bactericidal effects. It may react directly with proteins that contain transition metal centers, such as FeS, releasing Fe2+ for the Fenton reaction. Peroxynitrite may also react with various amino acids in the peptide chain, thereby altering protein structure and subsequently, protein function. It most commonly oxidises cysteine, and may indirectly induce tyrosine nitration through other generated RNS. Altered protein function includes changes in enzyme catalytic activity, cytoskeletal organisation and cell signal transduction.[4]
Hypochlorous acid reacts with a range of biomolecules, including DNA, lipids and proteins. HClO may oxidise cysteines and methionines via their sulfhydryl groups and sulfur groups respectively. The former leads to the formation of disulfide bonds, inducing protein crosslinking. Both oxidations result in protein aggregation, and ultimately, cell death.[5] Sulfhydryl groups can be oxidised up to three times by three HClO molecules, forming sulfenic acids, sulfinic acids and R–SO3H, which are increasingly irreversible and bactericidal.[6][7] Meanwhile, methionine oxidation is reversible. HOCl can also react with primary or secondary amines, producing chloroamines which are toxic to bacteria.[8][9] Protein cross linking and aggregation may also occur, as well as disruption of FeS groups.
Integral to hypochlorous acid formation is myeloperoxidase. Myeloperoxidase is most abundant in neutrophils, wherein phagocytosis is accompanied by degranulation. This is the fusion of granules with the phagolysosome, releasing their contents, including myeloperoxidase.[10] As many microbicidal products are formed during respiratory burst, the importance of individual molecules in killing invading pathogens is not wholly understood.
Due to the high toxicity of generated antimicrobial products including ROS, neutrophils have a short life span to limit host tissue damage during inflammation.
Disease
Chronic Granulomatous Disease is an inherited disease of human neutrophils, wherein NOX2 is defective. Phagocytosis may still occur, but without proper functioning NOX2, there is no superoxide production, and therefore no respiratory burst. The bacterial infection is not cleared.[11]
Cellular signalling
Non-phagocytic cells
In non-phagocytic cells, oxidative burst products are used in intracellular signalling pathways. The generated ROS achieve this via shifting the cell redox state. This may be monitored by the ratio of the antioxidant enzyme glutathione to its oxidised product, glutathione disulphide (GSH:GSSG).[12] Antioxidant enzymes counterbalance redox signalling by eliminating the involved molecules, importantly superoxide anion and nitric oxide. Redox signalling is critical for normal processes such as proliferation, differentiation, as well as vascular function and neurotransmission. It is also involved in disease states such as cancer.
The NADPH oxidase isoform NOX1 transiently produces a burst of superoxide in response to growth factor (e.g. EGF) stimulation of respective receptors.[13] Superoxide is dismutated to hydrogen peroxide at a rate close to the diffusion-limited rate. This spatial restriction for superoxide‘s dismutation allows for specificity of redox signalling. Specificity is also ensured by NOX1 localisation in specific microdomains in the cell’s plasma membrane. Through channels such as aquaporin or diffusion, hydrogen peroxide enters the cytosol. There, it oxidises the cysteine groups of redox-sensitive proteins, which can then transduce signals.[14]
Macrophages
Oxidative burst in phagocytes is most commonly associated with bacterial killing. However, macrophages, especially alveolar macrophages, usually produce far lower levels of ROS than neutrophils, and may require activation for their bactericidal properties. Instead, their transient oxidative burst regulates the inflammatory response by inducing cytokine synthesis for redox signalling, resulting in an influx of neutrophils and activated macrophages.[15]
Cancer cells
Cancer cells can manipulate cell signalling by producing excess levels of ROS, thereby constitutively activating pathways to promote their cellular growth and proliferation.[16] Implicated pathways include NF-κB, PI3K, HIFs and MAPKs. In humans, mitochondrial ROS is required alongside those released in the oxidative burst for mitogenic pathway stimulation in oncogenic KRAS cells. However, in oncogenic Kras mice fibroblasts, NADPH oxidase inhibitors have been shown to be sufficient to block these growth factor pathways.[17] Tumorigenic cells also simultaneously maintain high levels of antioxidants to protect against cancer cell death.[18]
Fertilisation
Most notably, oxidative burst post fertilisation can be seen in the sea urchin egg. This is believed to be evolutionally divergent from that in neutrophils.
Hydrogen peroxide is produced by egg oxidase activity following an increase in oxygen consumption.[19] This is essential for the cross-linking of the ovum proteins to prevent lethal polyspermy. Hydrogen peroxide itself is also spermicidal. However, the generated reactive species are maintained at lower levels than in immunity to protect the fertilised egg itself from oxidative damage. This is achieved by the elimination of hydrogen peroxide primarily through the dual function of the same egg oxidase, and secondarily through cytoplasmic ROS scavengers, such as catalase and glutathione.[20]
In plants
Oxidative burst acts as a defence mechanism to pathogen infection in plants. This is seen post PAMPs detection by cell-surface located receptors (e.g. FLS2 or EFR).[21][22][23] As in animals, the production of reactive oxygen species in plants is mediated by NADPH oxidase. In plant immunity, the NADPH oxidase subunits RbohD and RbohF have overlapping functions are expressed in different tissues and at different levels.[24][25] However, in contrast to animal phagocytes, wherein generated ROS are contained in the sealed phagolysosome, oxidative burst in plants is not contained. Consequently, generated ROS bear additional effects alongside pathogen toxicity. Hydrogen peroxide induces oxidative cross-linking of the plant’s cell wall glycoproteins.[26][27] This reduces susceptibility to enzymatic degradation by pathogens.[28] Systemic acquired resistance, which is analogous to innate immunity in animals, is also induced in the exposed plant cells.[29] Hydrogen peroxide exposure may also result in hypersensitive response, which is the death of a small number of host cells at the site of infection, for the purpose of limiting pathogenic infection.[30][31] ROS production in plants can be used as a readout for successful pathogen recognition via a luminol-peroxidase based assay.[32]
References
- Herb M, Schramm M (February 2021). "Functions of ROS in Macrophages and Antimicrobial Immunity". Antioxidants. 10 (2): 313. doi:10.3390/antiox10020313. PMC 7923022. PMID 33669824.
- Leto TL, Geiszt M (September 2006). "Role of Nox family NADPH oxidases in host defense". Antioxidants & Redox Signaling. 8 (9–10): 1549–61. doi:10.1089/ars.2006.8.1549. PMID 16987010.
- Imlay JA (2003). "Pathways of oxidative damage". Annual Review of Microbiology. 57: 395–418. doi:10.1146/annurev.micro.57.030502.090938. PMID 14527285.
- Pacher P, Beckman JS, Liaudet L (January 2007). "Nitric oxide and peroxynitrite in health and disease". Physiological Reviews. 87 (1): 315–424. doi:10.1152/physrev.00029.2006. PMC 2248324. PMID 17237348.
- Pereira WE, Hoyano Y, Summons RE, Bacon VA, Duffield AM (June 1973). "Chlorination studies. II. The reaction of aqueous hypochlorous acid with alpha-amino acids and dipeptides". Biochimica et Biophysica Acta. 313 (1): 170–80. doi:10.1016/0304-4165(73)90198-0. PMID 4745674.
- Winterbourn CC (June 1985). "Comparative reactivities of various biological compounds with myeloperoxidase-hydrogen peroxide-chloride, and similarity of the oxidant to hypochlorite". Biochimica et Biophysica Acta (BBA) - General Subjects. 840 (2): 204–10. doi:10.1016/0304-4165(85)90120-5. PMID 2986713.
- Prütz WA (January 1998). "Interactions of hypochlorous acid with pyrimidine nucleotides, and secondary reactions of chlorinated pyrimidines with GSH, NADH, and other substrates". Archives of Biochemistry and Biophysics. 349 (1): 183–91. doi:10.1006/abbi.1997.0440. PMID 9439597.
- Fang FC (6 September 2011). "Antimicrobial actions of reactive oxygen species". mBio. 2 (5). doi:10.1128/mBio.00141-11. PMC 3171981. PMID 21896680.
- Hampton MB, Kettle AJ, Winterbourn CC (November 1998). "Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing". Blood. 92 (9): 3007–17. doi:10.1182/blood.V92.9.3007.421k47_3007_3017. PMID 9787133.
- Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L (May 2000). "Neutrophils: molecules, functions and pathophysiological aspects". Laboratory Investigation; A Journal of Technical Methods and Pathology. 80 (5): 617–53. doi:10.1038/labinvest.3780067. PMID 10830774.
- Fernandez J. "Chronic Granulomatous Disease (CGD) - Immunology; Allergic Disorders". MSD Manual Professional Edition. Retrieved 2020-03-12.
- Schafer FQ, Buettner GR (June 2001). "Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple". Free Radical Biology & Medicine. 30 (11): 1191–212. doi:10.1016/S0891-5849(01)00480-4. PMID 11368918.
- Forman HJ, Torres M (December 2002). "Reactive oxygen species and cell signaling: respiratory burst in macrophage signaling". American Journal of Respiratory and Critical Care Medicine. 166 (12 Pt 2): S4-8. doi:10.1164/rccm.2206007. PMID 12471082.
- Di Marzo N, Chisci E, Giovannoni R (October 2018). "The Role of Hydrogen Peroxide in Redox-Dependent Signaling: Homeostatic and Pathological Responses in Mammalian Cells". Cells. 7 (10): 156. doi:10.3390/cells7100156. PMC 6211135. PMID 30287799.
- Forman HJ, Torres M (December 2002). "Reactive oxygen species and cell signaling: respiratory burst in macrophage signaling". American Journal of Respiratory and Critical Care Medicine. 166 (12 Pt 2): S4-8. doi:10.1164/rccm.2206007. PMID 12471082.
- Szatrowski TP, Nathan CF (February 1991). "Production of large amounts of hydrogen peroxide by human tumor cells". Cancer Research. 51 (3): 794–8. PMID 1846317.
- Irani K, Xia Y, Zweier JL, Sollott SJ, Der CJ, Fearon ER, et al. (March 1997). "Mitogenic signaling mediated by oxidants in Ras-transformed fibroblasts". Science. 275 (5306): 1649–52. doi:10.1126/science.275.5306.1649. PMID 9054359. S2CID 19733670.
- Gorrini C, Harris IS, Mak TW (December 2013). "Modulation of oxidative stress as an anticancer strategy". Nature Reviews. Drug Discovery. 12 (12): 931–47. doi:10.1038/nrd4002. PMID 24287781. S2CID 20604657.
- Warburg O (January 1908). "Beobachtungen über die Oxydationsprozesse im Seeigelei". Hoppe-Seyler's Zeitschrift für physiologische Chemie. 57 (1–2): 1–16. doi:10.1515/bchm2.1908.57.1-2.1.
- Wong JL, Créton R, Wessel GM (December 2004). "The oxidative burst at fertilization is dependent upon activation of the dual oxidase Udx1". Developmental Cell. 7 (6): 801–14. doi:10.1016/j.devcel.2004.10.014. PMID 15572124.
- Doke N (1985-11-01). "NADPH-dependent O2− generation in membrane fractions isolated from wounded potato tubers inoculated with Phytophthora infestans". Physiological Plant Pathology. 27 (3): 311–322. doi:10.1016/0048-4059(85)90044-X.
- Bradley DJ, Kjellbom P, Lamb CJ (July 1992). "Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: a novel, rapid defense response". Cell. 70 (1): 21–30. doi:10.1016/0092-8674(92)90530-P. PMID 1623521. S2CID 12312001.
- Jabs T, Tschope M, Colling C, Hahlbrock K, Scheel D (April 1997). "Elicitor-stimulated ion fluxes and O2- from the oxidative burst are essential components in triggering defense gene activation and phytoalexin synthesis in parsley". Proceedings of the National Academy of Sciences of the United States of America. 94 (9): 4800–5. Bibcode:1997PNAS...94.4800J. doi:10.1073/pnas.94.9.4800. PMC 20805. PMID 9114072.
- Torres MA, Dangl JL, Jones JD (January 2002). "Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response". Proceedings of the National Academy of Sciences of the United States of America. 99 (1): 517–22. Bibcode:2002PNAS...99..517T. doi:10.1073/pnas.012452499. PMC 117592. PMID 11756663.
- Morales J, Kadota Y, Zipfel C, Molina A, Torres MA (March 2016). "The Arabidopsis NADPH oxidases RbohD and RbohF display differential expression patterns and contributions during plant immunity". Journal of Experimental Botany. 67 (6): 1663–76. doi:10.1093/jxb/erv558. PMID 26798024.
- Bradley DJ, Kjellbom P, Lamb CJ (July 1992). "Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: a novel, rapid defense response". Cell. 70 (1): 21–30. doi:10.1016/0092-8674(92)90530-p. PMID 1623521. S2CID 12312001.
- Wojtaszek P, Trethowan J, Bolwell GP (September 1995). "Specificity in the immobilisation of cell wall proteins in response to different elicitor molecules in suspension-cultured cells of French bean (Phaseolus vulgaris L.)". Plant Molecular Biology. 28 (6): 1075–87. doi:10.1007/BF00032668. PMID 7548825. S2CID 23319754.
- Brisson LF, Tenhaken R, Lamb C (December 1994). "Function of Oxidative Cross-Linking of Cell Wall Structural Proteins in Plant Disease Resistance". The Plant Cell. 6 (12): 1703–1712. doi:10.1105/tpc.6.12.1703. PMC 160556. PMID 12244231.
- Chen Z, Silva H, Klessig DF (December 1993). "Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid". Science. 262 (5141): 1883–6. Bibcode:1993Sci...262.1883C. doi:10.1126/science.8266079. PMID 8266079.
- Tenhaken R, Levine A, Brisson LF, Dixon RA, Lamb C (May 1995). "Function of the oxidative burst in hypersensitive disease resistance". Proceedings of the National Academy of Sciences of the United States of America. 92 (10): 4158–63. Bibcode:1995PNAS...92.4158T. doi:10.1073/pnas.92.10.4158. PMC 41903. PMID 11607542.
- Levine A, Tenhaken R, Dixon R, Lamb C (November 1994). "H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response". Cell. 79 (4): 583–93. doi:10.1016/0092-8674(94)90544-4. PMID 7954825. S2CID 1488844.
- Keppler LD (1989). "Active Oxygen Production During a Bacteria-Induced Hypersensitive Reaction in Tobacco Suspension Cells". Phytopathology. 79 (9): 974. doi:10.1094/phyto-79-974.
External links
- Respiratory+burst at the US National Library of Medicine Medical Subject Headings (MeSH)