Pre-Bötzinger complex
The pre-Bötzinger complex (preBötC) is a cluster of interneurons in the ventral respiratory group of the medulla of the brainstem. This complex has been proven to be essential for the generation of the respiratory rhythm in mammals.[1] The exact mechanism of the rhythm generation and transmission to motor nuclei remains controversial and the topic of much research.[2][3][4][5][6][7]
| Pre-Bötzinger complex | |
|---|---|
| Identifiers | |
| NeuroLex ID | nlx_152600 |
| Anatomical terms of neuroanatomy | |
Several synthetic compounds have been shown to act on neurons specific to the preBötC, most being selective agonists or antagonists to receptor subtypes on neurons in the vicinity. Since many of these neurons express GABA, glutamate, serotonin[8] and adenosine receptors, chemicals custom tailored to bind at these sites are most effective at altering respiratory rhythm.
Adenosine modulates the preBötC output via activation of the A1 and A2A receptor subtypes.[9][10] An adenosine A1 receptor agonist has been shown to depress preBötC rhythmogenesis independent of the neurotransmitters GABA and glycine in in vitro preparations from 0- to 7-day-old mice.[11] Another synthetic drug specific to the adenosine A2A receptor subtype is CGS-21680 that has been shown to cause apneas in 14- to 21-day-old rat pups in vivo. For this reason, it has been used as a model to study pathological conditions such as apnea of prematurity and sudden infant death syndrome.
The region is named after the Silvaner (Boetzinger) variety of wine, served at the conference at which the region was named.
Description
The pre-Bötzinger complex (pre-BötC), a projection of the Botzinger complex, plays an important role in regulating respiration in mammals.[12] It is one of the four cell groups of the Ventral Respiratory Group (VRG). It is hypothesized that the pre-Bötzinger complex is the main location of the rhythmic pattern generation circuitry involved in breathing. It also contains cells that initiate spontaneous breathing. Research is being conducted on the mechanisms involved, but it is currently unclear how this interneuron system is regulated.
Rhythmogenesis, the generation of rhythm, is modulated by membrane properties and synaptic interactions occurring in interneurons, intrinsically bursting pacemaker neurons, and follower neurons within the pre-Bötzinger complex. Together these neurons make up an intrinsic network that is capable of being regulated by a vast range of neurotransmitters, amino acids, and chemical signals, such as adenosine, GABA, and glycine.[13]
The pre-Bötzinger complex produces two types of breathing rhythms under normal levels of oxygen. In eupnea, or normal breathing, the pre-BötC generates a rhythm that is fast and low in amplitude.[12] Sighs, on the other hand, consist of a slow and large amplitude rhythm. Each type of rhythm is generated by the same neurons in the pre-Bötzinger complex, but through different mechanisms, receptors, and ion currents that are controlled by changes in the behavior or environment of the organism. Under low levels of oxygen, the pre-Bötzinger complex needs to rearrange the activity of its neurons and requires the assistance of other brain structures, like the pons, to generate gasping. Gasping is characterized by a rhythm that has faster rise, shorter bursts, and lower frequency.
The pre-BötC is capable of generating stable respiratory rhythms with varying degrees of oxygenation and throughout varying conditions of its environment using network reconfiguration and neuromodulation.[12] The structure contains a core network within a larger network of interconnected nuclei that function to maintain respiratory rhythms and the neuroplasticity and state-dependency of breathing. Two key features of the pre-BötC are its stability and its ability to adapt to changing environmental and behavioral conditions.
Structure
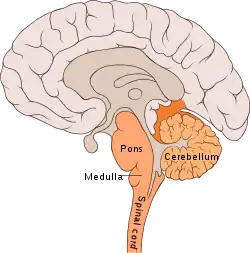
Located within the ventrolateral medulla, the pre-Bötzinger complex contains subnetworks that hold distinct synapses and intrinsic membrane properties.[12] In mammals, the respiratory network system and the nuclei controlling breathing modulation are found along the neuronal axis. The neuronal networks involved in respiratory function are located in the ventral respiratory column (VRC). From rostral to caudal, these networks include the retrotrapezoid nucleus/parafacial respiratory group complex (RTN/pFRG),[14] the Bötzinger complex, the pre-Bötzinger complex (pre-BötC), the rostral ventral respiratory group (rVRG), and the caudal VRG (cVRG). The dorsal pons, including the Kolliker-Fuse nucleus and the parabrachial complex, play an important role in respiratory control and rhythm generation. Other areas that aid in breathing control are the cerebellum, neocortex, and the periaqueductal gray (speech and breathing). The generation and modulation of eupnea and the transition between the inspiratory and expiratory phases have also been implicated in respiratory rhythm generation.
Important connections
The pre-BötC complex operates in animals as part of a larger network that receives critical information and signaling inputs from other areas of the brain, including the pons and parafacial nucleus.[12] An isolated slice of the complex can continue to generate respiratory rhythmic activity and can be recorded from the hypoglossal motor nucleus, which receives input through an outside interneuronal population.
Function
Types of breathing rhythms controlled
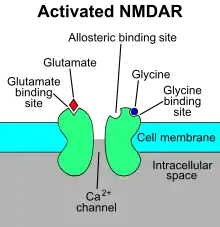
Rhythmic respiratory breathing continuously adapts to the organism's posture, activity level, speech, and can reveal whether someone is calm, agitated, or scared.[12] Plasticity of the mechanisms involved in respiratory behavior is modulated in part by the pre-Bötzinger complex. Disruption causes irreversible loss or major disruption of breathing in vivo. The frequency and amplitude change according to the behavioral and metabolic demands of the organism it controls. Breathing is thus extremely sensitive to the internal state of the organism. The pre-Bötzinger complex is a rhythm generating network, which is composed of micro networks that function within larger networks to create characteristic rhythms and breathing patterns, including eupneic, sigh, and gasping activities. This single neuronal network can create multiple respiratory rhythmic patterns and is by itself both necessary and sufficient to generate respiratory rhythms.
Its main function is to generate and coordinate respiratory rhythm and breathing cessation.[12] The complex also contributes to integrating modulatory and sensory inputs which feed its larger network with the capacity to adjust to changes in the behavioral, metabolic, and developmental features of the organism. Disturbances in neuromodulatory processes acting on ion channels, receptors, and second messengers have been associated with numerous pathophysiological conditions, such as Rett syndrome and sudden infant death syndrome.
Both the eupneic and the sigh rhythms involve activation of the majority of neurons. However, each type of rhythmic activity is dependent on distinct mechanisms. Fictive sigh depends critically on synaptic mechanisms that involve P/Q type calcium channels, suggestive of a subset of neurons with specialized synapses for this type of rhythm generation, since only a very small number of respiratory neurons receive glutamatergic inputs that depend on P/Q type calcium currents.[12] The fictive sigh rhythmic type also depends on mGluR8 receptor activation. Eupneic rhythmic activity relies on NMDA-dependent mechanisms.
When under a low oxygenated state (hypoxia) the respiratory network responds by transitioning into an augmentation followed by a depression phase, controlled in the pre-BötC.[12] During the depression phase, the inspiratory burst changes from an augmenting bell-shaped burst to a decrementing burst, a primary feature of gasping. Neuronal discharge patterns are altered during the depressed synaptic inhibition, contributing to the reformation of the network. Many of the respiratory neurons in the ventrolateral medulla inactivate before phrenic and/or hypoglossal (XII) cessation. These neurons are inconsistent in their response with rhythmic bursts and become either de- or hyperpolarized. A decrease in synaptic transmission is also seen with hypoxia.
Ionic currents

There are several inward currents that are proposed to help produce action potentials and bursts in pacemaker neurons. There are two main voltage dependent sodium currents that contribute to the depolarization and firing of action potentials in neurons. The fast and transient sodium current produces a large depolarization that fires the initial action potential in neurons, however this current is quickly inactivated and does not help maintain bursting activity in neurons.[13] To achieve bursts, a persistent sodium current provides enough depolarization to facilitate the firing of action potentials during a burst.[15] Unlike the fast and transient sodium current, the persistent sodium current (INaP) is activated at very low membrane potentials and has a much slower inactivation, which allows neurons to intrinsically fire action potentials at sub-threshold membrane potentials.[13] Studies have shown that the inactivation of this persistent sodium current helps end bursts in pacemaker neurons. The amount of time it takes for INaP to become activated again establishes the timeframe between each burst. The neuron can receive synaptic inputs and different amounts of inward and outward currents to regulate the time between each burst, which ultimately helps generate a specific breathing pattern.
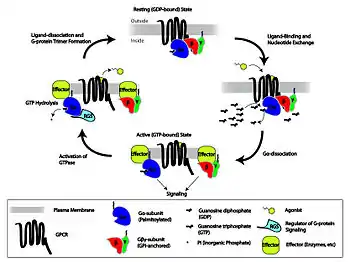
NALCN sodium leak channels have been hypothesized to give rise to an inward current that may play an important role in the modulation of bursting and spiking activity.[13] These nonselective cation channels may provide a voltage-independent sodium current that also helps slightly depolarize neurons. The channels are regulated by G protein–coupled receptors that can activate or inhibit the NALCN channels depending on the neurotransmitter that binds the receptor and the specific signaling pathway that is involved. Activation of M3 muscarinic receptors by acetylcholine and NK1 by Substance P significantly increases NALCN currents, while activation of CaSR by calcium stops the flow of the currents. Since NALCN sodium leak channels may contribute to the depolarization of neurons, their regulation by G-protein coupled receptors may be vital for the alteration of bursting and breathing rhythms.
Other inward currents that help generate intrinsic spiking and bursting in pacemaker neurons are the calcium current and calcium-activated nonspecific currents (ICAN).[13] When a neuron becomes depolarized, voltage gated calcium channels become activated and calcium is able to flow into the cell which usually leads to the release of neurotransmitters. Calcium-sensitive dyes have shown that internal concentrations of calcium increase during bursts. The activation of different calcium channels has distinct effects on the activity of neurons in the pre-Bötzinger complex. L-type calcium channels are known to increase the frequency of action potentials in some neurons, which might be the reason calcium influx through these channels has been observed during the augmentation when tissues have low levels of oxygen. P/Q-type calcium channels are mainly responsible for the release of neurotransmitters that excite, or activate, postsynaptic neurons. Studies have shown that blockage of these channels leads to the inhibition of sighs, which indicates calcium flow through these channels is necessary for sighs. Other research has also suggested that calcium flow through N-type calcium channels is essential for normal breathing, and is responsible for the activation of calcium-dependent potassium channels. Calcium-activated nonselective cation currents are important for the intrinsic spiking and bursting activity in CS pacemaker neurons. Metabotropic glutamate 1/5 receptors appear to be important for the increase in intracellular calcium that activate ICAN. The initial burst in a neuron usually leads to the activation of the transient sodium current and the several types of calcium currents. These currents depolarize the cell further enough to activate NMDA receptors and ICAN, which helps cell regenerate its bursts.
The ratio between inward and outward currents helps determine the activity of pacemaker neurons in the pre-Bötzinger complex. The major outward currents involved in the regulation of neuron activity are potassium currents.[13] Although the exact role of potassium currents is still being investigated, it appears that potassium and sodium leak currents are crucial for the rhythmicity of the pre-Bötzinger complex. Transient A-type potassium currents are more common in neurons that are involved in the inspiration process. When A-type potassium currents were blocked with 4-AP in slices of the pre-Bötzinger complex, synchronized bursts in inspiratory neurons was affected as well as communication with hypoglossal motor pools that help regulate breathing. This suggests that transient A-type potassium currents are needed for the synchronized bursts in inspiratory neurons and for effective respiratory control. Other potassium channels like large conductance calcium-dependent potassium channels and sodium chloride dependent potassium channels appear to end burst potentials in neurons. Moreover, ATP-dependent potassium channels help neurons detect changes in energy or oxygen levels to modify breathing patterns. These channels are activated by decreases in ATP, which suggests they provide the needed hyperpolarization during hypoxia.
Types of neurons
There are two types of neurons in the pre-BötC: nonpacemaker and pacemaker neurons. Nonpacemaker neurons enter either a tonic or a quiescent firing state, whereas the pacemaker neurons have spontaneous bursting potential.[12] Pacemaker neurons can further be subdivided into cadmium sensitive (CS) and cadmium insensitive (CI) pacemaker neurons. Bursting in CS neurons are mediated by ICAN, a nonspecific cation current, whereas CI is dependent upon the persistent sodium current, (INAP). Inhibiting these currents in the pacemaker subtypes cease their ability to spontaneously burst in synaptic isolation. Although they are a major contributor of inward current, these two currents are not the only ones mediating pacemakers. They also contribute to nonpacemaker neuronal properties.
The bursting properties of the pacemaker and nonpacemaker neuron run on a spectrum and have differing degrees of bursting potentials.[12] Neuromodulators, such as NE, SP, or 5-HT to name a few, can induce nonpacemaker bursting. Although the contributions of the nonpacemaker and pacemaker neurons are a gradient, they differ in their contribution when switching from eupneic to gasping activity. Eupneic activity requires the activation of ICAN and INaP-dependent bursting mechanisms, pacemaker neurons that on ICAN currents selectively hyperpolarize during low oxygenates states, meaning the system depend more so on INaP currents during gasping. The generation of sighs is more sensitive to INaP changes, therefore it may rely somewhat on pacemaker neuron contribution. Moreover, the complex respiratory network relies on different contributions of different types of bursting mechanisms.
Neuromodulation
The complex regulation of respiratory rhythm involves the integration of multiple signaling molecules and the activation of numerous diverse metabotropic and ionotropic receptors.[13] These include norepinephrine, serotonin, acetylcholine, substance P, ATP, TRH, somatostatin, dopamine, endorphins, and adenosine, which in turn activate g-protein coupled receptors to produce the diverse responses mediated by the pre-Bötzinger complex.
Nonpacemaker and pacemaker neurons involved in inspiration are stimulated by NE.[12] They are found within the pre-BötC and act via alpha-1, alpha-2, and beta-noradrenergic mechanisms. NE induces ICAN-dependent bursting in active nonpacemakers and depolarizes CI pacemakers, increasing the frequency of their bursting. In CS pacemakers, NE increases only the amplitude of the depolarizing drive potential and the number of action potentials during the burst,[12] but does not affect the burst frequency in CS pacemakers, unlike in CI pacemakers.
Serotonergic neurons are also involved in breathing systems.[12] Their actions are diverse and dependent upon the activity level and species of the animal. Serotonin plays a critical role in altering the pacemaker neurons involved in gasping and normal respiratory activity.[13] Blocking of the 5-HT2 receptor eliminates the bursts occurring in the pacemaker neurons and leads to the abolishing of gasps. The blocking of this receptor is therefore problematic, especially in SIDS, because gasping is an important mechanism involved in autoresuscitation. A lack of serotonin binding to the serotonin receptor 2 leads to the inability to autoresuscitation due to the lack of drive for gasping.
Substance P, a peptidergic modulator, also plays a role in neuromodulation of the pre-BötC.[12] It is often coreleased with other neurotransmitters. Substance P activates the inspiratory frequency at the level of the network and behavioral systems. Cellularly, substance P is involved in the depolarization of nonpacemaker neurons slowly, causing an increase in action potential firing rate. The neuropeptide can also activate CS pacemakers and less dramatically, CI pacemakers. This leads to an increase in burst amplitude, frequency, and duration. When Substance P is coreleased with serotonin, it plays a crucial role in hypoxic response.[13] This occurs because substance P stabilizes the respiratory rhythm through depolarization of neurons and activation of Pacemaker neurons.
Acetylcholine plays an important modulatory role on the respiratory system by altering nicotinic and muscarinic receptors.[13] The suppression of muscarinic receptors and the activation of nicotinic receptors due to prenatal exposure to nicotine have been linked to SIDS. This is due to the reduction of excitatory synaptic transmission in a nucleus and increased excitability in motor neurons caused by nicotinic activation.
Many other neuromodulators have roles in respiration. The aforementioned are simply three examples.
Homeostatic changes
Investigation of the respiratory response to Acute intermittent hypoxia (AIH), repeated episodes of hypoxia, reveals connection to various breathing disorders, such as Rett syndrome and obstructive sleep apnea.[12] AIH leads to persistent increases in respiratory frequency and amplitude of integrated motor neuronal bursts in vivo.[12] These changes lasting for 90 minutes or longer are termed long-term facilitation (LTF). AIH causes homeostatic changes in multiple sites of the respiratory system; the pre-BötC is likely the site for the LTF, since intermittent hypoxia causes an increase in persistent frequency after ongoing hypoxia. The respiratory system is regulated by multiple forms of long-term synaptic plasticity. The role of synaptic inhibition has been proved widespread and critical within the expiratory Botzinger complex respiratory network, through cross-correlation and antidromic mapping techniques. The inhibitory connections discovered indicate their ability to connect different classes of neurons, their importance in regulating the interval of inspiration, and their ability to control driving potential of respiratory neurons. These characteristics show the interaction between the parafacial respiratory group and the pre-Bötzinger complex, which allows for active expiration to be produced by synaptic inhibition within the respiratory network. Synaptic inhibition is critical for allowing the pre-Bötzinger complex to communicate with other respiratory centers in order to generate respiratory activity.
Glycinergic and GABAergic inhibitory neurons make up half of all inspiratory neurons. Exposure of the pre-Bötzinger complex to these inhibitory neurotransmitters results in the rhythmic nature associated with respiration. Blocking this inhibition from Glycine or GABA causes its neurons to be incapable of switching from the active phase to the inspiration phase, demonstrated by shorter inspiratory activity (as seen in vivo).[13] However, the absence of inhibitory synapses still resulted in rhythmic respiratory activity in vitro and in situ. This is largely due to the fact that respiratory rhythm results from numerous aspects, with synaptic inhibition playing only a single part.

In addition to the inhibitory synaptic regulation of respiratory rhythm within the pre-Bötzinger complex, there is also an excitatory component utilizing mostly AMPA receptors.[13] The generation of inspirations is due to a signaling cascade involving transient Ca2+ influx as a result of glutamate activating a postsynaptic receptor. In addition to glutamates role in activating the synaptic drive of inspiration, it is also understood that pacemaker neurons, with autonomous voltage-dependent properties, are also responsible for the generation of respiratory rhythm. Evidence of this is seen when isolating neurons within the pre-Bötzinger complex, which results in rhythmic bursts due to synaptically coupled micronetworks.
However, the generation of respiratory rhythm requires other excitatory components, such as glutamate, in order to produce a wide range of behavioral functions including eupneic and sigh activity.[13] The pre-Bötzinger complex is responsible for generating the wide variety of components that make up the respiratory rhythm. The accomplishment of these precise activities requires distinct neuron populations that overlap to allow the generation of different respiratory actions. Eupneic activity is generated using the excitatory mechanism through the NMDA glutamate receptor. Sighs have a differential generation originating from pacemaker neurons. The pre-Bötzinger complex is capable of generating differential rhythmic activities due to the intricate integration of modulatory, synaptic, and intrinsic properties of the neurons involved.
Oxygen sensing
In addition to its involvement in generating respiratory rhythm, the pre-Bötzinger complex is also capable of integrating sensory information from changes in the biochemical environment, particularly oxygen. The capability to detect focal hypoxia causes an excitatory response in the motor output responsible for respiration, which causes alterations in the firing pattern of neurons within the pre-Bötzinger complex.[13] Among these changes are the transition of a fully integrated network involving complex networks and autonomous mechanisms, to a system dependent on the activity of pacemaker neurons through sodium current activation. Hypoxia results in gasping due to the increased dependence on the sodium current and the overlap in networks between the generation of respiratory rhythm and intrinsic oxygen sensitization.
Associated diseases
References
- Smith JC, Ellenberger HH, Ballanyi K, Richter DW, Feldman JL (November 1991). "Pre-Bötzinger complex: a brainstem region that may generate respiratory rhythm in mammals". Science. 254 (5032): 726–9. doi:10.1126/science.1683005. PMC 3209964. PMID 1683005.
- Rybak IA, Abdala AP, Markin SN, Paton JF, Smith JC (2007). "Spatial organization and state-dependent mechanisms for respiratory rhythm and pattern generation". Computational Neuroscience: Theoretical Insights into Brain Function. Progress in Brain Research. Vol. 165. pp. 201–20. doi:10.1016/S0079-6123(06)65013-9. ISBN 978-0-444-52823-0. PMC 2408750. PMID 17925248.
- Smith JC, Abdala AP, Koizumi H, Rybak IA, Paton JF (December 2007). "Spatial and Functional Architecture of the Mammalian Brain Stem Respiratory Network: A Hierarchy of Three Oscillatory Mechanisms". Journal of Neurophysiology. 98 (6): 3370–87. doi:10.1152/jn.00985.2007. PMC 2225347. PMID 17913982.
- Gargaglioni LH, Bícegoa KC, Branco LG (December 2008). "Brain monoaminergic neurons and ventilatory control in vertebrates". Respiratory Physiology & Neurobiology. 164 (1–2): 112–22. doi:10.1016/j.resp.2008.04.017. PMID 18550453. S2CID 10661660.
- Rubin JE, Shevtsova NA, Ermentrout GB, Smith JC, Rybak IA (April 2009). "Multiple Rhythmic States in a Model of the Respiratory Central Pattern Generator". Journal of Neurophysiology. 101 (4): 2146–65. doi:10.1152/jn.90958.2008. PMC 2695631. PMID 19193773.
- Viemari JC, Tryba AK (April 2009). "Bioaminergic neuromodulation of respiratory rhythm in vitro". Respiratory Physiology & Neurobiology. 168 (1–2): 69–75. doi:10.1016/j.resp.2009.03.011. PMC 2791959. PMID 19538922.
- Abdala AP, Rybak IA, Smith JC, Zoccal DB, Machado BH, St-John WM, Paton JF (June 2009). "Multiple Pontomedullary Mechanisms of Respiratory Rhythmogenesis". Respiratory Physiology & Neurobiology. 168 (1–2): 19–25. doi:10.1016/j.resp.2009.06.011. PMC 2734878. PMID 19540366.
- Peña F, Ramirez JM (December 2002). "Endogenous activation of serotonin-2A receptors is required for respiratory rhythm generation in vitro". J. Neurosci. 22 (24): 11055–64. doi:10.1523/JNEUROSCI.22-24-11055.2002. PMC 6758407. PMID 12486201.
- Mayer CA, Haxhiu MA, Martin RJ, Wilson CG (January 2006). "Adenosine A2A receptors mediate GABAergic inhibition of respiration in immature rats". Journal of Applied Physiology. 100 (1): 91–7. doi:10.1152/japplphysiol.00459.2005. PMID 16141383.
- Vandam RJ, Shields EJ, Kelty JD (2008). "Rhythm generation by the pre-Bötzinger Complex in medullary slice and island preparations: Effects of adenosine A1 receptor activation". BMC Neuroscience. 9: 95. doi:10.1186/1471-2202-9-95. PMC 2567986. PMID 18826652.
- Kuwana S, Tsunekawa N, Yanagawa Y, Okada Y, Kuribayashi J, Obata K (February 2006). "Electrophysiological and morphological characteristics of GABAergic respiratory neurons in the mouse pre-Bötzinger complex". The European Journal of Neuroscience. 23 (3): 667–74. doi:10.1111/j.1460-9568.2006.04591.x. PMID 16487148. S2CID 21509969.
- Garcia AJ, 3rd; Zanella, S; Koch, H; Doi, A; Ramirez, JM (2011). Chapter 3--networks within networks: the neuronal control of breathing. Progress in Brain Research. Vol. 188. pp. 31–50. doi:10.1016/B978-0-444-53825-3.00008-5. ISBN 9780444538253. PMC 3652403. PMID 21333801.
- Ramirez JM, Doi A, Garcia AJ, Elsen FP, Koch H, Wei AD (2012). "The Cellular Building Blocks of Breathing". The cellular building blocks of breathing. Comprehensive Physiology. Vol. 2. pp. 2683–2731. doi:10.1002/cphy.c110033. ISBN 9780470650714. PMC 3684023. PMID 23720262.
- Guyenet, PG; Stornetta, RL; Abbott, SB; Depuy, SD; Kanbar, R (2012). The retrotrapezoid nucleus and breathing. Advances in Experimental Medicine and Biology. Vol. 758. pp. 115–22. doi:10.1007/978-94-007-4584-1_16. ISBN 978-94-007-4583-4. PMC 5111164. PMID 23080151.
- Stafstrom CE (2007). "Persistent Sodium Current and Its Role in Epilepsy". Epilepsy Currents. 7 (1): 15–22. doi:10.1111/j.1535-7511.2007.00156.x. PMC 1797888. PMID 17304346.