Aposematism
Aposematism is the advertising by an animal to potential predators that it is not worth attacking nor eating.[1] This unprofitability may consist of any defences which make the prey difficult to kill and eat, such as toxicity, venom, foul taste or smell, sharp spines, or aggressive nature. These advertising signals may take the form of conspicuous coloration, sounds, odours,[2] or other perceivable characteristics. Aposematic signals are beneficial for both predator and prey, since both avoid potential harm.


The term was coined in 1877 by Edward Bagnall Poulton[3][4] for Alfred Russel Wallace's concept of warning coloration.[5] Aposematism is exploited in Müllerian mimicry, where species with strong defences evolve to resemble one another. By mimicking similarly coloured species, the warning signal to predators is shared, causing them to learn more quickly at less of a cost.
A genuine aposematic signal that a species actually possesses chemical or physical defences is not the only way to deter predators. In Batesian mimicry, a mimicking species resembles an aposematic model closely enough to share the protection, while many species have bluffing deimatic displays which may startle a predator long enough to enable an otherwise undefended prey to escape.
There is good evidence for aposematism in terrestrial animals; its existence in marine animals is possible but disputed.
Etymology
The term aposematism was coined by the English zoologist Edward Bagnall Poulton in his 1890 book The Colours of Animals. He based the term on the Ancient Greek words ἀπό apo 'away' and σῆμα sēma 'sign', referring to signs that warn other animals away.[3][4]
Defence mechanism

The function of aposematism is to prevent attack, by warning potential predators that the prey animal has defences such as being unpalatable or poisonous. The easily detected warning is a primary defence mechanism, and the non-visible defences are secondary.[6] Aposematic signals are primarily visual, using bright colours and high-contrast patterns such as stripes. Warning signals are honest indications of noxious prey, because conspicuousness evolves in tandem with noxiousness.[7] Thus, the brighter and more conspicuous the organism, the more toxic it usually is.[7][8] This is in contrast to deimatic displays, which attempt to startle a predator with a threatening appearance but which are bluffing, unsupported by any strong defences.[9]
The most common and effective colours are red, yellow, black, and white.[10] These colours provide strong contrast with green foliage, resist changes in shadow and lighting, are highly chromatic, and provide distance dependent camouflage.[10] Some forms of warning coloration provide this distance dependent camouflage by having an effective pattern and colour combination that do not allow for easy detection by a predator from a distance, but are warning-like from a close proximity, allowing for an advantageous balance between camouflage and aposematism.[11] Warning coloration evolves in response to background, light conditions, and predator vision.[12] Visible signals may be accompanied by odours, sounds or behaviour to provide a multi-modal signal which is more effectively detected by predators.[13]

Unpalatability, broadly understood, can be created in a variety of ways. Some insects such as the ladybird or tiger moth contain bitter-tasting chemicals,[14] while the skunk produces a noxious odour, and the poison glands of the poison dart frog, the sting of a velvet ant or neurotoxin in a black widow spider make them dangerous or painful to attack. Tiger moths advertise their unpalatability by either producing ultrasonic noises which warn bats to avoid them,[14] or by warning postures which expose brightly coloured body parts (see Unkenreflex), or exposing eyespots. Velvet ants (actually parasitic wasps) such as Dasymutilla occidentalis both have bright colours and produce audible noises when grabbed (via stridulation), which serve to reinforce the warning.[15] Among mammals, predators can be dissuaded when a smaller animal is aggressive and able to defend itself, as for example in honey badgers.[16]
Prevalence
In terrestrial ecosystems

Aposematism is widespread in insects, but less so in vertebrates, being mostly confined to a smaller number of reptile, amphibian, and fish species, and some foul-smelling or aggressive mammals. Pitohuis, red and black birds whose toxic feathers and skin apparently comes from the poisonous beetles they ingest, could be included.[17] It has been proposed that aposematism played a role in human evolution, body odour carrying a warning to predators of large hominins able to defend themselves with weapons.[18]
Perhaps the most numerous aposematic vertebrates are the poison dart frogs (family: Dendrobatidae).[19] These neotropical anuran amphibians exhibit a wide spectrum of coloration and toxicity.[20] Some species in this poison frog family (particularly Dendrobates, Epipedobates, and Phyllobates) are conspicuously coloured and sequester one of the most toxic alkaloids among all living species.[21][22] Within the same family, there are also cryptic frogs (such as Colostethus and Mannophryne) that lack these toxic alkaloids.[23][24] Although these frogs display an extensive array of coloration and toxicity, there is very little genetic difference between the species.[20] Evolution of their conspicuous coloration is correlated to traits such as chemical defense, dietary specialization, acoustic diversification, and increased body mass.[25][22]
Some plants are thought to employ aposematism to warn herbivores of unpalatable chemicals or physical defences such as prickled leaves or thorns.[26] Many insects, such as cinnabar moth caterpillars, acquire toxic chemicals from their host plants.[27] Among mammals, skunks and zorillas advertise their foul-smelling chemical defences with sharply contrasting black-and-white patterns on their fur, while the similarly-patterned badger and honey badger advertise their sharp claws, powerful jaws, and aggressive natures.[28] Some brightly coloured birds such as passerines with contrasting patterns may also be aposematic, at least in females; but since male birds are often brightly coloured through sexual selection, and their coloration is not correlated with edibility, it is unclear whether aposematism is significant.[29]
The sound-producing rattle of rattlesnakes is an acoustic form of aposematism.[30] Sound production by the caterpillar of the Polyphemus moth, Antheraea polyphemus, may similarly be acoustic aposematism, connected to and preceded by chemical defences.[31] Similar acoustic defences exist in a range of Bombycoidea caterpillars.[32]
In marine ecosystems


The existence of aposematism in marine ecosystems is controversial.[35] Many marine organisms, particularly those on coral reefs, are brightly coloured or patterned, including sponges, corals, molluscs and fish, with little or no connection to chemical or physical defenses. Caribbean reef sponges are brightly coloured, and many species are full of toxic chemicals, but there is no statistical relationship between the two factors.[36]
Nudibranch molluscs are the most commonly cited examples of aposematism in marine ecosystems, but the evidence for this has been contested,[37] mostly because (1) there are few examples of mimicry among species, (2) many species are nocturnal or cryptic, and (3) bright colours at the red end of the colour spectrum are rapidly attenuated as a function of water depth. For example, the Spanish Dancer nudibranch (genus Hexabranchus), among the largest of tropical marine slugs, potently chemically defended, and brilliantly red and white, is nocturnal and has no known mimics. Mimicry is to be expected as Batesian mimics with weak defences can gain a measure of protection from their resemblance to aposematic species.[38] Other studies have concluded that nudibranchs such as the slugs of the family Phyllidiidae from Indo-Pacific coral reefs are aposematically coloured.[39] Müllerian mimicry has been implicated in the coloration of some Mediterranean nudibranchs, all of which derive defensive chemicals from their sponge diet.[40]
The crown-of-thorns starfish, like other starfish such as Metrodira subulata, has conspicuous coloration and conspicuous long, sharp spines, as well as cytolytic saponins, chemicals which could function as an effective defence; this evidence is argued to be sufficient for such species to be considered aposematic.[33][34] It has been proposed that aposematism and mimicry is less evident in marine invertebrates than terrestrial insects because predation is a more intense selective force for many insects, which disperse as adults rather than as larvae and have much shorter generation times.[35] Further, there is evidence that fish predators such as blueheads may adapt to visual cues more rapidly than do birds, making aposematism less effective.[41]
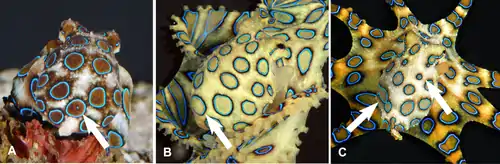
Blue-ringed octopuses are venomous. They spend much of their time hiding in crevices whilst displaying effective camouflage patterns with their dermal chromatophore cells. However, if they are provoked, they quickly change colour, becoming bright yellow with each of the 50-60 rings flashing bright iridescent blue within a third of a second.[42] It is often stated this is an aposematic warning display,[43][44][45][46] but the hypothesis has rarely if ever been tested.[47]
Behaviour
The mechanism of defence relies on the memory of the would-be predator; a bird that has once experienced a foul-tasting grasshopper will endeavour to avoid a repetition of the experience. As a consequence, aposematic species are often gregarious. Before the memory of a bad experience attenuates, the predator may have the experience reinforced through repetition. Aposematic organisms often move in a languid fashion, as they have little need for speed and agility. Instead, their morphology is frequently tough and resistant to injury, thereby allowing them to escape once the predator is warned off. Aposematic species do not need to hide or stay still as cryptic organisms do, so aposematic individuals benefit from more freedom in exposed areas and can spend more time foraging, allowing them to find more and better quality food.[48] They may also be able to make use of conspicuous mating displays, including vocal signals, which may then develop through sexual selection.[49][22]
Origins of the theory

Wallace, 1867
In a letter to Alfred Russel Wallace dated 23 February 1867, Charles Darwin wrote, "On Monday evening I called on Bates & put a difficulty before him, which he could not answer, & as on some former similar occasion, his first suggestion was, 'you had better ask Wallace'. My difficulty is, why are caterpillars sometimes so beautifully & artistically coloured?"[50] Darwin was puzzled because his theory of sexual selection (where females choose their mates based on how attractive they are) could not apply to caterpillars since they are immature and hence not sexually active.
Wallace replied the next day with the suggestion that since some caterpillars "...are protected by a disagreeable taste or odour, it would be a positive advantage to them never to be mistaken for any of the palatable catterpillars [sic], because a slight wound such as would be caused by a peck of a bird’s bill almost always I believe kills a growing catterpillar. Any gaudy & conspicuous colour therefore, that would plainly distinguish them from the brown & green eatable catterpillars, would enable birds to recognise them easily as at a kind not fit for food, & thus they would escape seizure which is as bad as being eaten."[51]
Since Darwin was enthusiastic about the idea, Wallace asked the Entomological Society of London to test the hypothesis.[52] In response, the entomologist John Jenner Weir conducted experiments with caterpillars and birds in his aviary, and in 1869 he provided the first experimental evidence for warning coloration in animals.[53] The evolution of aposematism surprised 19th-century naturalists because the probability of its establishment in a population was presumed to be low, since a conspicuous signal suggested a higher chance of predation.[54]
Poulton, 1890
.jpg.webp)
Wallace coined the term "warning colours" in an article about animal coloration in 1877.[5] In 1890 Edward Bagnall Poulton renamed the concept aposematism in his book The Colours of Animals.[4] He described the derivation of the term as follows:
The second head (Sematic Colours) includes Warning Colours and Recognition Markings: the former warn an enemy off, and are therefore called Aposematic [Greek, apo, from, and sema, sign][55]
Evolution
Aposematism is paradoxical in evolutionary terms, as it makes individuals conspicuous to predators, so they may be killed and the trait eliminated before predators learn to avoid it.[56] If warning coloration puts the first few individuals at such a strong disadvantage, it would never last in the species long enough to become beneficial.[57]
Supported explanations
There is evidence for explanations involving dietary conservatism, in which predators avoid new prey because it is an unknown quantity;[58] this is a long-lasting effect.[58][59][60] Dietary conservatism has been demonstrated experimentally in some species of birds and fish.[61][58][60][62]
Further, birds recall and avoid objects that are both conspicuous and foul-tasting longer than objects that are equally foul-tasting but cryptically coloured.[63] This suggests that Wallace's original view, that warning coloration helped to teach predators to avoid prey thus coloured, was correct.[64] However, some birds (inexperienced starlings and domestic chicks) also innately avoid conspicuously coloured objects, as demonstrated using mealworms painted yellow and black to resemble wasps, with dull green controls. This implies that warning coloration works at least in part by stimulating the evolution of predators to encode the meaning of the warning signal, rather than by requiring each new generation to learn the signal's meaning.[64] All of these results contradict the idea that novel, brightly coloured individuals would be more likely to be eaten or attacked by predators.[58][65]
Alternative hypotheses
Other explanations are possible. Predators might innately fear unfamiliar forms (neophobia)[66] long enough for them to become established, but this is likely to be only temporary.[57][66][67]
Alternatively, prey animals might be sufficiently gregarious to form clusters tight enough to enhance the warning signal. If the species was already unpalatable, predators might learn to avoid the cluster, protecting gregarious individuals with the new aposematic trait.[68][69] Gregariousness would assist predators to learn to avoid unpalatable, gregarious prey.[70] Aposematism could also be favoured in dense populations even if these are not gregarious.[58][66]
Another possibility is that a gene for aposematism might be recessive and located on the X chromosome.[71] If so, predators would learn to associate the colour with unpalatability from males with the trait, while heterozygous females carry the trait until it becomes common and predators understand the signal.[71] Well-fed predators might also ignore aposematic morphs, preferring other prey species.[57][72]
A further explanation is that females might prefer brighter males, so sexual selection could result in aposematic males having higher reproductive success than non-aposematic males if they can survive long enough to mate. Sexual selection is strong enough to allow seemingly maladaptive traits to persist despite other factors working against the trait.[19]
Once aposematic individuals reach a certain threshold population, for whatever reason, the predator learning process would be spread out over a larger number of individuals and therefore is less likely to wipe out the trait for warning coloration completely.[73] If the population of aposematic individuals all originated from the same few individuals, the predator learning process would result in a stronger warning signal for surviving kin, resulting in higher inclusive fitness for the dead or injured individuals through kin selection.[74]
A theory for the evolution of aposematism posits that it arises by reciprocal selection between predators and prey, where distinctive features in prey, which could be visual or chemical, are selected by non-discriminating predators, and where, concurrently, avoidance of distinctive prey is selected by predators. Concurrent reciprocal selection (CRS) may entail learning by predators or it may give rise to unlearned avoidances by them. Aposematism arising by CRS operates without special conditions of the gregariousness or the relatedness of prey, and it is not contingent upon predator sampling of prey to learn that aposematic cues are associated with unpalatability or other unprofitable features.[75]
Mimicry
Aposematism is a sufficiently successful strategy to have had significant effects on the evolution of both aposematic and non-aposematic species.
Non-aposematic species have often evolved to mimic the conspicuous markings of their aposematic counterparts. For example, the hornet moth is a deceptive mimic of the yellowjacket wasp; it resembles the wasp, but has no sting. A predator which avoids the wasp will to some degree also avoid the moth. This is known as Batesian mimicry, after Henry Walter Bates, a British naturalist who studied Amazonian butterflies in the second half of the 19th century.[76] Batesian mimicry is frequency dependent: it is most effective when the ratio of mimic to model is low; otherwise, predators will encounter the mimic too often.[77][78]
A second form of mimicry occurs when two aposematic organisms share the same anti-predator adaptation and non-deceptively mimic each other, to the benefit of both species, since fewer individuals of either species need to be attacked for predators to learn to avoid both of them. This form of mimicry is known as Müllerian mimicry, after Fritz Müller, a German naturalist who studied the phenomenon in the Amazon in the late 19th century.[79][80] Many species of bee and wasp that occur together are Müllerian mimics; their similar coloration teaches predators that a striped pattern is associated with being stung. Therefore, a predator which has had a negative experience with any such species will likely avoid any that resemble it in the future. Müllerian mimicry is found in vertebrates such as the mimic poison frog (Ranitomeya imitator) which has several morphs throughout its natural geographical range, each of which looks very similar to a different species of poison frog which lives in that area.[81]
 A model (to be mimicked), the venomous and genuinely aposematic coral snake
A model (to be mimicked), the venomous and genuinely aposematic coral snake The harmless red milk snake, a Batesian mimic of the coral snake
The harmless red milk snake, a Batesian mimic of the coral snake
See also
- Handicap principle
References
- Santos, J. C.; Coloma, Luis A.; Cannatella, D. C. (2003). "Multiple, recurring origins of aposematism and diet specialization in poison frogs". Proceedings of the National Academy of Sciences. 100 (22): 12792–12797. doi:10.1073/pnas.2133521100. PMC 240697. PMID 14555763.
- Eisner, T.; Grant, R. P. (1981). "Toxicity, Odor Aversion, and 'Olfactory Aposematism'". Science. 213 (4506): 476. Bibcode:1981Sci...213..476E. doi:10.1126/science.7244647. PMID 7244647.
- Poulton 1890, pp. Foldout "The Colours of Animals Classified According to Their Uses", after page 339.
- Marek, Paul. "Aposematism". Apheloria. Archived from the original on 8 July 2017. Retrieved November 24, 2012.
- Wallace, Alfred Russel (1877). "The Colours of Animals and Plants. I.—The Colours of Animals". Macmillan's Magazine. 36 (215): 384–408.
- Ruxton, Sherratt & Speed 2004, pp. 82–103.
- Maan, M. E.; Cummings, M. E. (2012). "Poison frog colors are honest signals of toxicity, particularly for bird predators". American Naturalist. 179 (1): E1–E14. doi:10.1086/663197. hdl:2152/31175. JSTOR 663197. PMID 22173468. S2CID 1963316.
- Blount, Jonathan D.; Speed, Michael P.; Ruxton, Graeme D.; et al. (2009). "Warning displays may function as honest signals of toxicity". Proceedings of the Royal Society B. 276 (1658): 871–877. doi:10.1098/rspb.2008.1407. PMC 2664363. PMID 19019790.
- Sendova-Franks, Ana; Scott, Michelle Pellissier (2015). "Featured Articles in This Month's Animal Behaviour". Animal Behaviour. 100: iii–v. doi:10.1016/j.anbehav.2014.12.013. S2CID 53202282.
- Stevens, M.; Ruxton, G. D. (2012). "Linking the evolution and form of warning coloration in nature". Proceedings of the Royal Society B: Biological Sciences. 279 (1728): 417–426. doi:10.1098/rspb.2011.1932. PMC 3234570. PMID 22113031.
- Tullberg, B. S., Merilaita S., & Wiklund, C. (2005). "Aposematism and Crypsis Combined as a Result of Distance Dependence: Functional Versatility of the Colour Pattern in the Swallowtail Butterfly Larva". Proceedings: Biological Sciences. 272 (1570): 1315–1321. doi:10.1098/rspb.2005.3079. PMC 1560331. PMID 16006332.
{{cite journal}}: CS1 maint: uses authors parameter (link) - Wang, I. J.; Shaffer, H. B. (2008). "Rapid color evolution in an aposematic species: A phylogenetic analysis of colour variation in the strikingly polymorphic strawberry poison-dart frog". Evolution. 62 (11): 2742–2759. doi:10.1111/j.1558-5646.2008.00507.x. PMID 18764916.
- MacAuslane, Heather J. (2008). "Aposematism". In Capinera (ed.). Encyclopedia Entomologica. Vol. 4. pp. 239–242.
- Hristov, N. I.; Conner, W. E. (2005). "Sound strategy: acoustic aposematism in the bat–tiger moth arms race". Naturwissenschaften. 92 (4): 164–169. Bibcode:2005NW.....92..164H. doi:10.1007/s00114-005-0611-7. PMID 15772807. S2CID 18306198.
- Schmidt, J. O.; Blum, M. S. (1977). "Adaptations and Responses of Dasymutilla occidentalis (Hymenoptera: Mutillidae) to Predators". Entomologia Experimentalis et Applicata. 21 (2): 99–111. doi:10.1111/j.1570-7458.1977.tb02663.x. S2CID 83847876.
- "Black, White and Stinky: Explaining Coloration in Skunks and Other Boldly Colored Animals". University of Massachusetts Amherst. 27 May 2011. Retrieved 21 March 2016.
- Dumbacher, J. P.; Beehler, B. M.; Spande, T. F.; et al. (1992). "Homobatrachotoxin in the genus Pitohui: chemical defense in birds?". Science. 258 (5083): 799–801. Bibcode:1992Sci...258..799D. doi:10.1126/science.1439786. PMID 1439786.
- Weldon, Paul J. (18 August 2018). "Are we chemically aposematic? Revisiting L. S. B. Leakey's hypothesis on human body odour". Biological Journal of the Linnean Society. 125 (2): 221–228. doi:10.1093/biolinnean/bly109.
- Maan, M. E.; Cummings, M. E. (2009). "Sexual dimorphism and directional sexual selection on aposematic signals in a poison frog". Proceedings of the National Academy of Sciences of the United States of America. 106 (45): 19072–19077. Bibcode:2009PNAS..10619072M. doi:10.1073/pnas.0903327106. PMC 2776464. PMID 19858491.
- Summers, Kyle; Clough, Mark E. (2001-05-22). "The evolution of coloration and toxicity in the poison frog family (Dendrobatidae)". Proceedings of the National Academy of Sciences. 98 (11): 6227–6232. Bibcode:2001PNAS...98.6227S. doi:10.1073/pnas.101134898. PMC 33450. PMID 11353830.
- Santos, Juan C.; Tarvin, Rebecca D.; O’Connell, Lauren A. (2016). Schulte, Bruce A.; Goodwin, Thomas E.; Ferkin, Michael H. (eds.). "A Review of Chemical Defense in Poison Frogs (Dendrobatidae): Ecology, Pharmacokinetics, and Autoresistance". Chemical Signals in Vertebrates. Springer International Publishing. 13: 305–337. doi:10.1007/978-3-319-22026-0_21. ISBN 9783319220260. S2CID 85845409.
- Santos, Juan C.; Baquero, Margarita; Barrio-Amorós, César; et al. (2014-12-07). "Aposematism increases acoustic diversification and speciation in poison frogs". Proceedings of the Royal Society B: Biological Sciences. 281 (1796): 20141761. doi:10.1098/rspb.2014.1761. PMC 4213648. PMID 25320164.
- Guillory, Wilson X.; Muell, Morgan R.; Summers, Kyle; et al. (August 2019). "Phylogenomic Reconstruction of the Neotropical Poison Frogs (Dendrobatidae) and Their Conservation". Diversity. 11 (8): 126. doi:10.3390/d11080126.
- "Multiple, recurring origins of aposematism and diet specialization in poison frogs (PDF Download Available)". ResearchGate. Retrieved 2017-11-11.
- Santos, Juan C.; Cannatella, David C. (2011-04-12). "Phenotypic integration emerges from aposematism and scale in poison frogs". Proceedings of the National Academy of Sciences. 108 (15): 6175–6180. Bibcode:2011PNAS..108.6175S. doi:10.1073/pnas.1010952108. PMC 3076872. PMID 21444790.
- Rubino, Darrin L.; McCarthy, Brian C. "Presence of Aposematic (Warning) Coloration in Vascular Plants of Southeastern Ohio" Journal of the Torrey Botanical Society, Vol. 131, No. 3 (Jul-Sep 2004), pp. 252-256. https://www.jstor.org/stable/4126955
- Edmunds 1974, pp. 199–201.
- Caro, Tim (2009). "Contrasting coloration in terrestrial mammals". Philosophical Transactions of the Royal Society B: Biological Sciences. 364 (1516): 537–548. doi:10.1098/rstb.2008.0221. PMC 2674080. PMID 18990666.
- Götmark, Frank (1994). "Are Bright Birds Distasteful? A Re-Analysis of H. B. Cott's Data on the Edibility of Birds". Journal of Avian Biology. 25 (3): 184–197. doi:10.2307/3677074. JSTOR 3677074.
- Meik, Jesse M.; Pires-daSilva, André (10 February 2009). "Evolutionary morphology of the rattlesnake style". BMC Evolutionary Biology. 9 (1): 35. doi:10.1186/1471-2148-9-35. PMC 2645363. PMID 19208237.
- Brown, Sarah G.; Boettner, George H.; Yack, Jayne E. (15 March 2007). "Clicking caterpillars: acoustic aposematism in Antheraea polyphemus and other Bombycoidea". The Journal of Experimental Biology. 210 (6): 993–1005. doi:10.1242/jeb.001990. PMID 17337712.
- Bura, Veronica L.; Kawahara, Akito Y.; Yack, Jayne E. (11 August 2016). "A Comparative Analysis of Sonic Defences in Bombycoidea Caterpillars". Scientific Reports. 6 (1): 31469. Bibcode:2016NatSR...631469B. doi:10.1038/srep31469. PMC 4980592. PMID 27510510.
- Shedd, John G. (2006). "Crown of Thorns Sea Star". Shedd Aquarium. Archived from the original on 22 February 2014. Retrieved 21 June 2015.
- Inbar, Moshe; Lev-Yadun, Simcha (2005). "Conspicuous and aposematic spines in the animal kingdom" (PDF). Naturwissenschaften. 92 (4): 170–172. Bibcode:2005NW.....92..170I. doi:10.1007/s00114-005-0608-2. PMID 15761732. S2CID 8525349.
defensive animal spines are often conspicuous (shape and colour) and should be considered aposematic... Classic examples are the starfishes Acanthaster planci and Metrodira subulata, which have red spines...
- Pawlik, J. R. (2012). Fattorusso, E.; et al. (eds.). Antipredatory defensive roles of natural products from marine invertebrates. Handbook of Marine Natural Products. Springer. pp. 677–710.
- Pawlik, J. R.; et al. (1995). "Defenses of Caribbean sponges against predatory reef fish: I. Chemical deterrenc". Marine Ecology Progress Series. 127: 183–194. Bibcode:1995MEPS..127..183P. doi:10.3354/meps127183.
- Edmunds, Malcolm (1991). "Does warning coloration occur in nudibranchs?". Malacologia. 32: 241–255.
- Pawlik, J. R.; et al. (1988). "Defensive chemicals of the Spanish Dancer nudibranch, Hexabranchus sanguineus, and its egg ribbons: Macrolides derived from a sponge diet". Journal of Experimental Marine Biology and Ecology. 119 (2): 99–109. doi:10.1016/0022-0981(88)90225-0.
- Ritson-Williams, R.; Paul, V. J. (2007). "Marine benthic invertebrates use multimodal cues for defense against reef fish". Marine Ecology Progress Series. 340: 29–39. Bibcode:2007MEPS..340...29R. doi:10.3354/meps340029.
- Haber, M.; et al. (2010). "Coloration and defense in the nudibranch gastropod Hypselodoris fontandraui". Biological Bulletin. 218 (2): 181–188. doi:10.1086/BBLv218n2p181. PMID 20413794. S2CID 10948319.
- Miller, A. M.; Pawlik, J. R. (2013). "Do coral reef fish learn to avoid unpalatable prey using visual cues?". Animal Behaviour. 85 (2): 339–347. doi:10.1016/j.anbehav.2012.11.002. S2CID 43941481.
- Mäthger, L. M. (2012). Bell, G. R., Kuzirian, A. M., Allen, J. J. and Hanlon, R. T. "How does the blue-ringed octopus (Hapalochlaena lunulata) flash its blue rings?". The Journal of Experimental Biology. 215 (21): 3752–3757. doi:10.1242/jeb.076869. PMID 23053367.
- Williams, B. L. (2011). Lovenburg, V., Huffard, C. L. and Caldwell, R. L. "Chemical defense in pelagic octopus paralarvae: Tetrodotoxin alone does not protect individual paralarvae of the greater blue-ringed octopus (Hapalochlaena lunulata) from common reef predators". Chemoecology. 21 (3): 131–141. doi:10.1007/s00049-011-0075-5. S2CID 9953958.
- Huffard, C. L., Saarman, N., Hamilton, H. and Simison, W. B. (2010). "The evolution of conspicuous facultative mimicry in octopuses: an example of secondary adaptation?". Biological Journal of the Linnean Society. 101 (1): 68–77. doi:10.1111/j.1095-8312.2010.01484.x.
{{cite journal}}: CS1 maint: uses authors parameter (link) - Lambert, W. A. (2011). A Review of Blue-ringed Octopus Conservation (Masters thesis). Prescott College.
- Hanlon, R. T.; Messenger, J. B. (1998). Cephalopod Behaviour. Cambridge University Press.
- Umbers, K. D. (2013). "On the perception, production and function of blue coloration in animals". Journal of Zoology. 289 (4): 229–242. doi:10.1111/jzo.12001.
- Speed, M. P.; Brockhurst, M. A.; Ruxton, G. D. (2010). "The dual benefits of aposematism: predator avoidance and enhanced resources collection". Evolution. 64 (6): 1622–1633. doi:10.1111/j.1558-5646.2009.00931.x. PMID 20050915. S2CID 21509940.
- Rudh, A.; Rogell, B.; Håstad, O.; et al. (2011). "Rapid population divergence linked with co-variation between coloration and sexual display in strawberry poison frogs". Evolution. 65 (5): 1271–1282. doi:10.1111/j.1558-5646.2010.01210.x. PMID 21166789. S2CID 10785432.
- Darwin, Charles. "Letter from Charles Robert Darwin to Alfred Russel Wallace dated 23 February [1867]".
- Wallace, Alfred Russel. "Letter from Alfred Russel Wallace to Charles Robert Darwin dated 24 February [1867]".
- Anon (1867). "Discussion [Wallace's explanation of brilliant colors in caterpillar larvae, and others' comments thereon, presented at the ESL meeting of 4 March 1867]". Journal of Proceedings of the Entomological Society of London: lxxx–lxxxi.
- Slotten, Ross (2004). The Heretic in Darwin's Court:The Life of Alfred Russel Wallace. New York: Columbia University Press. pp. 253–524. ISBN 978-0-231-13010-3.
- Higashi, Masahiko; Yachi, Shigeo (August 1998). "The evolution of warning signals". Nature. 394 (6696): 882–884. Bibcode:1998Natur.394..882Y. doi:10.1038/29751. S2CID 204999972.
- Poulton 1890, pp. 337–338.
- Briolat, Emmanuelle S.; Burdfield-Steel, Emily R.; Paul, Sarah C.; et al. (2019). "Diversity in warning coloration: selective paradox or the norm?". Biological Reviews. 94 (2): 388–414. doi:10.1111/brv.12460. PMC 6446817. PMID 30152037.
- Mappes, Johanna; Marples, Nicola; Endler, John A. (2005). "The complex business of survival by aposematism". Trends in Ecology and Evolution. 20 (11): 598–603. doi:10.1016/j.tree.2005.07.011. PMID 16701442.
- Thomas, R. J.; Marples, N. M.; Cuthill, Innes C.; et al. (2003). "Dietary conservatism may facilitate the initial evolution of aposematism". Oikos. 101 (3): 548–566. doi:10.1034/j.1600-0706.2003.12061.x.
- Marples, Nicola M.; Kelly, David J.; Thomas, Robert J. (2005). "Perspective: The evolution of warning coloration is not paradoxical". Evolution. 59 (5): 933–940. doi:10.1111/j.0014-3820.2005.tb01032.x. PMID 16136793.
- Lindstrom, Leena; Altalo, Rauno V.; Lyytinen, Anne; et al. (2001). "Predator experience on cryptic prey affects the survival of conspicuous aposematic prey". Proceedings of the Royal Society. 268 (1465): 357–361. doi:10.1098/rspb.2000.1377. PMC 1088614. PMID 11270431.
- Richards, E. Loys; Thomas, Robert J.; Marples, Nicola M.; et al. (2011). "The expression of dietary conservatism in solitary and shoaling 3-spined sticklebacks Gasterosteus aculeatus". Behavioral Ecology. 22 (4): 738–744. doi:10.1093/beheco/arr047.
- Richards, E. Loys; Alexander, Lucille G.; Snellgrove, Donna; et al. (February 2014). "Variation in the expression of dietary conservatism within and between fish species". Animal Behaviour. 88: 49–56. doi:10.1016/j.anbehav.2013.11.009. S2CID 53174146.
- Siddall, Emma C.; Marples, Nicola M. (2008-01-22). "Better to be bimodal: the interaction of color and odor on learning and memory". Behavioral Ecology. 19 (2): 425–432. doi:10.1093/beheco/arm155.
- Roper, Tim J. (9 July 1987). "All things bright and poisonous". New Scientist. Reed Business Information. pp. 50–52.
- Marples, Nicola M.; Kelly, David J.; Thomas, Robert J. (May 2005). "Perspective: The Evolution of Warning Coloration is Not Paradoxical". Evolution. 59 (5): 933–940. doi:10.1111/j.0014-3820.2005.tb01032.x. PMID 16136793.
- Speed, Michael P. (2001). "Can receiver psychology explain the evolution of aposematism?". Animal Behaviour. 61 (1): 205–216. doi:10.1006/anbe.2000.1558. PMID 11170710. S2CID 13434024.
- Exernova, Alice; Stys, Pavel; Fucikova, Eva; et al. (2007). "Avoidance of aposematic prey in European tits (Paridae): learned or innate?". Behavioral Ecology. 18 (1): 148–156. doi:10.1093/beheco/arl061.
- Gamberale, Gabriella; Tullberg, Birgitta S. (1998). "Aposematism and gregariousness: the combined effect of group size and coloration on signal repellence". Proceedings of the Royal Society B: Biological Sciences. 265 (1399): 889–894. doi:10.1098/rspb.1998.0374. PMC 1689056.
- Mappes, Johanna; Alatalo, Rauno V. (1996). "Effects of novelty and gregariousness in survival of aposematic prey". Behavioral Ecology. 8 (2): 174–177. doi:10.1093/beheco/8.2.174.
- Ruxton, Graeme D.; Sherratt, Thomas N. (2006). "Aggregation, defense, and warning signals: the evolutionary relationship". Proceedings of the Royal Society. 273 (1600): 2417–2424. doi:10.1098/rspb.2006.3570. PMC 1634906. PMID 16959629.
- Brodie, Edmund D. III; Agrawal, Anell F. (2001). "Maternal effects and the evolution of aposematic signals". Proceedings of the National Academy of Sciences. 98 (14): 7884–7887. Bibcode:2001PNAS...98.7884B. doi:10.1073/pnas.141075998. PMC 35437. PMID 11416165.
- Merilaita, Sami; Kaitala, Veijo (2002). "Community structure and the evolution of aposematic coloration". Ecology Letters. 5 (4): 495–501. doi:10.1046/j.1461-0248.2002.00362.x. S2CID 7188102.
- Lee, T. J.; Marples, N. M.; Speed, M. P. (2010). "Can dietary conservatism explain the primary evolution of aposematism?". Animal Behaviour. 79: 63–74. doi:10.1016/j.anbehav.2009.10.004. S2CID 54273453.
- Servedio, M. R. (2000). "The effects of predator learning, forgetting, and recognition errors on the evolution of warning coloration". Evolution. 54 (3): 751–763. doi:10.1111/j.0014-3820.2000.tb00077.x. PMID 10937250.
- Weldon, P.J. (December 2013). "Chemical aposematism". Chemoecology. 23 (4): 201–202. doi:10.1007/s00049-013-0140-3. S2CID 27025142.
- Bates, Henry Walter (1861). "Contributions to an insect fauna of the Amazon valley. Lepidoptera: Heliconidae". Transactions of the Linnean Society. 23 (3): 495–566. doi:10.1111/j.1096-3642.1860.tb00146.x.; Reprint: Bates, Henry Walter (1981). "Contributions to an insect fauna of the Amazon valley (Lepidoptera: Heliconidae)". Biological Journal of the Linnean Society. 16 (1): 41–54. doi:10.1111/j.1095-8312.1981.tb01842.x.
- Harper, G. R.; Pfennig, D. W (22 August 2007). "Mimicry on the edge: why do mimics vary in resemblance to their model in different parts of their geographical range?". Proceedings of the Royal Society B: Biological Sciences. 274 (1621): 1955–1961. doi:10.1098/rspb.2007.0558. PMC 2275182. PMID 17567563.
- Edmunds 1974, p. 112.
- Müller, Fritz (1878). "Ueber die Vortheile der Mimicry bei Schmetterlingen" [On the advantages of mimicry in butterflies]. Zoologischer Anzeiger (in German). 1: 54–55.
- Müller, Fritz (1879). Translated by R. Meldola. "Ituna and Thyridia; a remarkable case of mimicry in butterflies". Proclamations of the Entomological Society of London. 1879: 20–29.
- Twomey, Evan; Brown, Jason (1986). "Ranitomeya imitator". Dendrobates.org. Retrieved 11 May 2015.
Sources
- Edmunds, Malcolm (1974). Defence in Animals. Longman. ISBN 978-0-582-44132-3.
- Poulton, Edward Bagnall (1890). The Colours of Animals, their meaning and use, especially considered in the case of insects. London: Kegan Paul, Trench & Trübner.
- Ruxton, Graeme D.; Sherratt, T. N.; Speed, M. P. (2004). Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals and Mimicry. Oxford University Press. ISBN 978-0-19-852859-3.
External links
 Media related to Warning coloration at Wikimedia Commons
Media related to Warning coloration at Wikimedia Commons
Vision in animals | ||
|---|---|---|
| Vision |
| .jpg.webp) |
| Eyes |
| |
| Evolution |
| |
| Coloration |
| |
| Related topics |
| |