Daspletosaurus
Daspletosaurus (/dæsˌpliːtəˈsɔːrəs/ das-PLEET-ə-SOR-əs; meaning "frightful lizard") is a genus of tyrannosaurid dinosaur that lived in Laramidia between about 79.5 and 74 million years ago, during the Late Cretaceous Period. The genus Daspletosaurus contains two species. Fossils of the earlier type species, D. torosus, have been found in Alberta, while fossils of the later second species, D. horneri, have been found only in Montana. A possible third species, also from Alberta, awaits formal identification and another possible species D. degrootorum, also exists, but it may belong to the separate genus Thanatotheristes instead. Daspletosaurus is closely related to the much larger and more recent tyrannosaurid Tyrannosaurus rex. Like most tyrannosaurids, Daspletosaurus was a multi-tonne bipedal predator equipped with dozens of large, sharp teeth. Daspletosaurus had the small forelimbs typical of tyrannosaurids, although they were proportionately longer than in other genera.
| Daspletosaurus Temporal range: Late Cretaceous (Campanian), | |
|---|---|
 | |
| Cast skeleton mount of D. torosus on display at Milwaukee Public Museum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Tyrannosauridae |
| Clade: | †Daspletosaurini |
| Genus: | †Daspletosaurus Russell, 1970 |
| Type species | |
| †Daspletosaurus torosus Russell, 1970 | |
| Other species | |
| |
| Synonyms | |
| |
As an apex predator, Daspletosaurus was at the top of the food chain, probably preying on large dinosaurs like the ceratopsid Centrosaurus and the hadrosaur Hypacrosaurus. In some areas, Daspletosaurus coexisted with another tyrannosaurid, Gorgosaurus, though there is some evidence of niche differentiation between the two.[2] While Daspletosaurus fossils are not as common as other tyrannosaurid fossils, the available specimens allow some analysis of the biology of these animals, including social behavior, diet, and life history.[3]
Discovery and naming

The type specimen of Daspletosaurus torosus (CMN 8506) is a partial skeleton including the skull, the shoulder, a forelimb, the pelvis, a femur, and all of the vertebrae from the neck, torso, and hip, as well as the first eleven tail vertebrae. It was discovered in 1921 near Steveville, Alberta, by Charles Mortram Sternberg, who thought it was a new species of Gorgosaurus. It was not until 1970 that the specimen was fully described by Dale Russell, who made it the type of a new genus, Daspletosaurus, from the Greek δασπλής (dasplēs, stem and connective vowel resulting in dasplēto-) ("frightful") and σαυρος (sauros) ("lizard").[4] The type species is Daspletosaurus torosus, the specific name torosus being Latin for 'muscular' or 'brawny'.[5] Aside from the type, there is only one other well-known specimen, RTMP 2001.36.1, a relatively complete skeleton discovered in 2001. Both specimens were recovered from the Oldman Formation in the Judith River Group of Alberta. The Oldman Formation was deposited during the middle Campanian stage of the Late Cretaceous, from about 79.5 to 77 Ma (million years ago).[6]
Dale Russell also suggested that a specimen of an immature Albertosaurus (CMN 11315) from the younger Horseshoe Canyon Formation in Alberta actually belonged to a third specimen of Daspletosaurus as D. cf. torosus, extending the temporal range of the genus by approximately 3.5 million years into the Maastrichtian. He based this referral on features of its limb and pelvic girdle, as well as the curvature of the hand claws, which he interpreted as traits matching Daspletosaurus. This reassignment was not universally accepted, and thorough re-examination of the specimen favoured its initial referral to Albertosaurus sarcophagus, despite lacking many of the diagnostic skeletal traits used to identify mature tyrannosaurids.[7][8] An additional maxilla and various teeth from an Edmontosaurus-dominated bonebed in the Horseshoe Canyon Formation was also mistakenly referred to Daspletosaurus, but all the tyrannosaurid material has all since been confirmed to belong to Albertosaurus.[9]
Assigned species

Two or three additional species have been assigned to the genus Daspletosaurus over the years, although as of 2007 none of these species has received a proper description or scientific name. In the meantime, all are designated as Daspletosaurus spp; this does not imply that they all are the same species.[7][10]

Along with the holotype, Russell designated a specimen collected by Barnum Brown in 1913 as the paratype of D. torosus. This specimen (AMNH 5438) consists of parts of the hindleg, the pelvis, and some of its associated vertebrae. It was discovered in the Dinosaur Park Formation in Alberta.[7] The Dinosaur Park Formation was formerly known as the Upper Oldman Formation and dates back to the middle Campanian, between 76.5 and 74.8 million years ago.[11] Daspletosaurus fossils are known specifically from the middle to upper section of the formation, between 75.6 and 75.0 million years ago.[12] In 1914, Brown collected a nearly complete skeleton and skull; forty years later his American Museum of Natural History sold this specimen to the Field Museum of Natural History in Chicago. It was mounted for display in Chicago and labeled as Albertosaurus libratus for many years, but after several skull features were later found to be modeled in plaster, including most of the teeth, the specimen (FMNH PR308) was reassigned to Daspletosaurus torosus by Thomas Carr in 1999.[13] A total of eight specimens have been collected from the Dinosaur Park Formation over the years since, most of them within the boundaries of Dinosaur Provincial Park. Phil Currie believes that the Dinosaur Park specimens represent a new species of Daspletosaurus, distinguished by certain features of the skull. Pictures of this new species have been published, but it still awaits a name and full description in print.[7]
A new tyrannosaurid specimen (OMNH 10131), including skull fragments, ribs, and parts of the hindlimb, was reported from New Mexico in 1990 and assigned to the now-defunct genus Aublysodon.[14] Many later authors have reassigned this specimen, along with a few others from New Mexico, to yet another unnamed species of Daspletosaurus.[7][10][15] However, research published in 2010 showed that this species, from the Hunter Wash Member of the Kirtland Formation, is actually a more primitive tyrannosauroid, and was classified in the genus Bistahieversor.[16]

In 1992, Jack Horner and colleagues published an extremely preliminary report of a tyrannosaurid from the upper parts of the Campanian Two Medicine Formation in Montana, which was interpreted as a transitional species between Daspletosaurus and the later Tyrannosaurus.[17] Currie (2003) stated that the tyrannosaurid from the Two Medicine Formation mentioned by Horner et al. (1992) may be an unnamed third species of Daspletosaurus.[7] Another partial skeleton was reported from the Upper Two Medicine in 2001, preserving the remains of a juvenile hadrosaur in its abdominal cavity. This specimen was assigned to Daspletosaurus but not to any particular species.[18] The remains of at least three more Daspletosaurus have also been described in a Two Medicine bonebed by Currie et al. (2005); the authors stated that this fossil material likely represents then-unnamed species mentioned by Horner et al. (1992), but cautioned that further study and description of Daspletosaurus would be necessary before the species can be determined with certainty.[19] In 2017, the Two Medicine Formation taxon was named as the new species D. horneri.[20]
Isolated tyrannosaurid teeth in the upper portions of the Judith River Formation are likely from Gorgosaurus as well as some species of Daspletosaurus, probably D. torosus. However, in the lower portion Judith River formation, around 78 million years ago, there is some evidence for a new undescribed tyrannosaurid taxon. A specimen in the collections of Triebold Paleontology excavated between 2002 and 2004, known as "Sir William", shows some characteristics of Daspletosaurus suggesting a new earlier species to the genus. However the specimen shows many characteristics typical of early tyrannosaurines such as Teratophoneus and even some of the later Tyrannosaurus, which may suggest an entirely new genus.[21]
Description
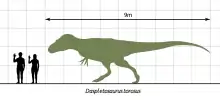
While very large by the standard of modern predators, Daspletosaurus was not the largest tyrannosaurid. Adults could reach a length of 8–9 meters (26–30 ft) from snout to tail,[5] and a body mass of 2–3 metric tons (2.2–3.3 short tons).[22][5][23][24][25]

Daspletosaurus had a massive skull that could reach more than 1 meter (3 ft 3 in) in length.[5] The bones were heavily constructed and some, including the nasal bones on top of the snout, were fused for strength. Large fenestrae (openings) in the skull reduced its weight. An adult Daspletosaurus was armed with about six dozen teeth that were very long but oval in cross section rather than blade-like. Unlike its other teeth, those in the premaxilla at the end of the upper jaw had a D-shaped cross section, an example of heterodonty always seen in tyrannosaurids. Unique skull features included the rough outer surface of the maxilla (upper jaw bone) and the pronounced crests around the eyes on the lacrimal, postorbital, and jugal bones. The orbit (eye socket) was a tall oval, somewhere in between the circular shape seen in Gorgosaurus and the 'keyhole' shape of Tyrannosaurus.[13][7][10] Split carinae (edges)[26] have been found on Daspletosaurus teeth.[27]
Daspletosaurus shared the same body form as other tyrannosaurids, with a short, S-shaped neck supporting the massive skull. It walked on its two thick hindlimbs, which ended in four-toed feet, although the first digit (the hallux) did not contact the ground. In contrast, the forelimbs were extremely small and bore only two digits, although Daspletosaurus had the longest forelimbs in proportion to body size of any tyrannosaurid. A long, heavy tail served as a counterweight to the head and torso, with the center of gravity over the hips.[5][10]
Classification and systematics

Daspletosaurus belongs in the subfamily Tyrannosaurinae within the family Tyrannosauridae, along with Tarbosaurus, Tyrannosaurus, and Alioramus. Animals in this subfamily are more closely related to Tyrannosaurus than to Albertosaurus and are known – with the exception of Alioramus – for their robust build with proportionally larger skulls and longer femora than in the other subfamily, the Albertosaurinae.[10][28] It further belongs to the tribe Daspletosaurini, consisting of it and the taxon Thanatotheristes.[29]
Daspletosaurus is usually considered to be closely related to Tyrannosaurus rex, or even a direct ancestor through anagenesis.[17] Gregory Paul reassigned D. torosus to the genus Tyrannosaurus, creating the new combination Tyrannosaurus torosus,[23] but this has not been generally accepted.[13][10] Many researchers believe Tarbosaurus and Tyrannosaurus to be sister taxa or even to be the same genus, with Daspletosaurus a more basal relative.[10][30] On the other hand, Phil Currie and colleagues find Daspletosaurus to be more closely related to Tarbosaurus and other Asian tyrannosaurids like Alioramus than to the North American Tyrannosaurus.[28] The systematics (evolutionary relationships) of Daspletosaurus may become clearer once all the species have been described.



Below is the cladogram of Tyrannosauridae based on the phylogenetic analysis conducted by Loewen et al. in 2013.[31]
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
Senses
There are indications of D. horneri possessing integumentary sensory organs, possibly used in touch, modulation of precise jaw movements, temperature reading, and prey detection. The large flat scales may have further protected the snout during prey capture and intra-specific combat.[32][33][20]
Social behavior

A young specimen of the Dinosaur Park Daspletosaurus species (TMP 94.143.1) shows bite marks on the face that were inflicted by another tyrannosaur. The bite marks are healed over, indicating that the animal survived the bite. A full-grown Dinosaur Park Daspletosaurus (TMP 85.62.1) also exhibits tyrannosaur bite marks, showing that attacks to the face were not limited to younger animals. While it is possible that the bites were attributable to other species, intraspecific aggression, including facial biting, is very common among predators. Facial bites are seen in other tyrannosaurs like Gorgosaurus and Tyrannosaurus, as well as in other theropod genera like Sinraptor and Saurornitholestes. Darren Tanke and Phil Currie hypothesize that the bites are due to intraspecific competition for territory or resources, or for dominance within a social group.[34]
Evidence that Daspletosaurus lived in social groups comes from a bonebed found in the Two Medicine Formation of Montana. The bonebed includes the remains of three Daspletosaurus, including a large adult, a small juvenile, and another individual of intermediate size. At least five hadrosaurs are preserved at the same location. Geologic evidence indicates that the remains were not brought together by river currents but that all of the animals were buried simultaneously at the same location. The hadrosaur remains are scattered and bear numerous marks from tyrannosaur teeth, indicating that the Daspletosaurus were feeding on the hadrosaurs at the time of death. The cause of death is unknown. Currie speculates that the daspletosaurs formed a pack, although this cannot be stated with certainty.[19] Other scientists are skeptical of the evidence for social groups in Daspletosaurus and other large theropods;[35]

Brian Roach and Daniel Brinkman have suggested that Daspletosaurus social interaction would have more closely resembled the modern Komodo dragon, where non-cooperative individuals mob carcasses, frequently attacking and even cannibalizing each other in the process.[36] Evidence of cannibalism in Daspletosaurus was published in 2015.[37]
Life history
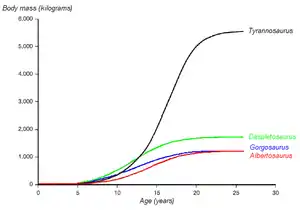
Paleontologist Gregory Erickson and colleagues have studied the growth and life history of tyrannosaurids. Analysis of bone histology can determine the age of a specimen when it died. Growth rates can be examined when the ages of various individuals are plotted against their size on a graph. Erickson has shown that after a long time as juveniles, tyrannosaurs underwent tremendous growth spurts for about four years midway through their lives. After the rapid growth phase ended with sexual maturity, growth slowed down considerably in adult animals. Erickson only examined Daspletosaurus from the Dinosaur Park Formation, but these specimens show the same pattern. Compared to albertosaurines, Daspletosaurus showed a faster growth rate during the rapid growth period due to its higher adult weight. The maximum growth rate in Daspletosaurus was 180 kilograms (400 lb) per year, based on a mass estimate of 1,800 kilograms (2.0 short tons) in adults. Other authors have suggested higher adult weights for Daspletosaurus; this would change the magnitude of the growth rate but not the overall pattern.[38]

By tabulating the number of specimens of each age group, Erickson and his colleagues were able to draw conclusions about life history in a population of Albertosaurus. Their analysis showed that, while juveniles were rare in the fossil record, subadults in the rapid growth phase and adults were far more common. While this could be due to preservation or collection biases, Erickson hypothesized that the difference was due to low mortality among juveniles over a certain size, which is also seen in some modern large mammals like elephants. This low mortality may have resulted from a lack of predation, since tyrannosaurs surpassed all contemporaneous predators in size by the age of two. Paleontologists have not found enough Daspletosaurus remains for a similar analysis, but Erickson notes that the same general trend seems to apply.[39]
A 2009 study found evidence of Trichomonas gallinae-like infection in the jaws of various specimens of Daspletosaurus.[40]
Paleoecology

All known Daspletosaurus fossils have been found in formations dating to the middle to late Campanian stage of the Late Cretaceous Period, between 77 and 74 million years ago. Since the middle of the Cretaceous, North America had been divided in half by the Western Interior Seaway, with much of Montana and Alberta below the surface. However, the uplift of the Rocky Mountains in the Laramide Orogeny to the west, which began during the time of Daspletosaurus, forced the seaway to retreat eastwards and southwards. Rivers flowed down from the mountains and drained into the seaway, carrying sediment along with them that formed the Two Medicine Formation, the Judith River Group, and other sedimentary formations in the region. About 73 million years ago, the seaway began to advance westwards and northwards again, and the entire region was covered by the Bearpaw Sea, represented throughout the western United States and Canada by the massive Bearpaw Shale.[41][42][43]
Daspletosaurus lived in a vast floodplain along the western shore of the interior seaway. Large rivers watered the land, occasionally flooding and blanketing the region with new sediment. When water was plentiful, the region could support a great deal of plant and animal life, but periodic droughts also struck the region, resulting in mass mortality as preserved in the many bonebed deposits found in Two Medicine and Judith River sediments, including the Daspletosaurus bonebed.[44] Similar conditions exist today in East Africa.[45] Volcanic eruptions from the west periodically blanketed the region with ash, also resulting in large-scale mortality, while simultaneously enriching the soil for future plant growth. It is these ash beds that allow precise radiometric dating as well. Fluctuating sea levels also resulted in a variety of other environments at different times and places within the Judith River Group, including offshore and nearshore marine habitats, coastal wetlands, deltas, and lagoons, in addition to the inland floodplains.[42] The Two Medicine Formation was deposited at higher elevations farther inland than the other two formations.[43]
The excellent vertebrate fossil record of Two Medicine and Judith River rocks resulted from a combination of abundant animal life, periodic natural disasters, and the deposition of large amounts of sediment. Many types of freshwater and estuarine fish are represented, including sharks, rays, sturgeons, gars, and others. The Judith River Group preserves the remains of many aquatic amphibians and reptiles, including frogs, salamanders, turtles, Champsosaurus and crocodilians. Terrestrial lizards, including whiptails, skinks, monitors, and alligator lizards have also been discovered. Azhdarchid pterosaurs, and birds like Apatornis and Avisaurus flew overhead, while several varieties of mammals coexisted with Daspletosaurus and other types of dinosaurs in the various formations that make up the Judith River wedge.[42]

In the Oldman Formation (the geological equivalent of the Judith River formation), Daspletosaurus torosus could have preyed upon the hadrosaur species Brachylophosaurus canadensis, the ceratopsians Coronosaurus brinkmani and Albertaceratops nesmoi, pachycephalosaurs, ornithomimids, therizinosaurs, and possibly ankylosaurs. Other predators included troodontids, oviraptorosaurs, the dromaeosaurid Saurornitholestes, and possibly an albertosaurine tyrannosaur (genus currently unknown). The younger Dinosaur Park and Two Medicine Formations had faunas similar to the Oldman, with the Dinosaur Park in particular preserving an unrivaled array of dinosaurs.[42] The albertosaurine Gorgosaurus lived alongside unnamed species of Daspletosaurus in the Dinosaur Park and Upper Two Medicine environments.[46] Young tyrannosaurs may have filled the niches in between adult tyrannosaurs and smaller theropods, which were separated by two orders of magnitude in mass.[5][10][47][48] A Saurornitholestes dentary has been discovered in the Dinosaur Park Formation that bore tooth marks left by the bite of a young tyrannosaur, possibly Daspletosaurus.[49]
Coexistence with Gorgosaurus

In the late Campanian of North America, Daspletosaurus was a contemporary of the albertosaurine tyrannosaurid Gorgosaurus. This is one of the few examples of two tyrannosaur genera coexisting. In modern predator guilds, similar-sized predators are separated into different ecological niches by anatomical, behavioral or geographical differences that limit competition.[46] Several studies have attempted to explain niche differentiation in Daspletosaurus and Gorgosaurus.
Dale Russell hypothesized that the more lightly built and more common Gorgosaurus may have preyed on the abundant hadrosaurs of the time, while the more robust and less common Daspletosaurus may have specialized on the less prevalent but better-defended ceratopsids, which may have been more difficult to hunt.[5] However, a specimen of Daspletosaurus (OTM 200) from the Two Medicine Formation preserves the digested remains of a juvenile hadrosaur in its gut region.[18] The higher and broader muzzles of tyrannosaurines like Daspletosaurus are mechanically stronger than the lower snouts of albertosaurines like Gorgosaurus, although tooth strengths are similar between the two groups. This may indicate a difference in feeding mechanics or diet.[47]
Other authors have suggested that competition was limited by geographical separation. Unlike some other groups of dinosaurs, there appears to be no correlation with distance from the sea. Neither Daspletosaurus nor Gorgosaurus was more common at higher or lower elevations than the other.[46] However, while there is some overlap, Gorgosaurus appears to be more common at northern latitudes, with species of Daspletosaurus more abundant to the south. The same pattern is seen in other groups of dinosaurs. Chasmosaurine ceratopsians and hadrosaurine hadrosaurs (a group now generally referred to as saurolophines) are also more common in the Two Medicine Formation and in southwestern North America during the Campanian. Thomas Holtz has suggested that this pattern indicates shared ecological preferences between tyrannosaurines, chasmosaurines and hadrosaurines. Holtz notes that, at the end of the later Maastrichtian stage, tyrannosaurines like Tyrannosaurus rex, hadrosaurines and chasmosaurines like Triceratops were widespread throughout western North America, while albertosaurines and centrosaurines became extinct, and lambeosaurines were very rare.[10]
See also
- Timeline of tyrannosaur research
- 2017 in archosaur paleontology
References
- Chan-gyu Yun (2020). "A Subadult Frontal of Daspletosaurus torosus (Theropoda: Tyrannosauridae) from the Late Cretaceous of Alberta, Canada with Implications for Tyrannosaurid Ontogeny and Taxonomy". PalArch's Journal of Vertebrate Palaeontology. 17: 1–13. Archived from the original on September 27, 2020. Retrieved January 15, 2021.
- Yun, Changyu (2021). Tyrannosaurid theropod specimens in the San Diego Natural History Museum from the Dinosaur Park Formation (Campanian) of Alberta, Canada. New Mexico Museum of Natural History and Science. pp. 569–578.
- Trexler, David; Currie, Phillip; Koppelhus, Eva; Wicks, Kelly; Murphy, Nate (June 2005). Carpenter, Kenneth (ed.). The Carnivorous Dinosaurs. Bloomington, Ind: Indiana University Press. pp. 313–324. ISBN 9780253345394. Retrieved January 12, 2022.
- Liddell, Henry G.; Scott, Robert (1980). Greek-English Lexicon (Abridged ed.). Oxford: Oxford University Press. ISBN 978-0-19-910207-5.
- Russell, Dale A. (1970). "Tyrannosaurs from the Late Cretaceous of western Canada". National Museum of Natural Sciences Publications in Paleontology. 1: 1–34.
- Fowler, Denver Warwick (November 22, 2017). "Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian-Maastrichtian (Late Cretaceous) formations of the Western Interior of North America". PLOS ONE. 12 (11): e0188426. Bibcode:2017PLoSO..1288426F. doi:10.1371/journal.pone.0188426. ISSN 1932-6203. PMC 5699823. PMID 29166406.
- Currie, Philip J. (2003). "Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta" (PDF). Acta Palaeontologica Polonica. 48 (2): 191–226.
- Mallon, J.C.; Bura, J.R.; Currie, P.J. (2019). "A Problematic Tyrannosaurid (Dinosauria: Theropoda) Skeleton and Its Implications for Tyrannosaurid Diversity in the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta". The Anatomical Record. 303 (4): 673–690. doi:10.1002/ar.24199. PMC 7079176. PMID 31254458.
- Torices, A.; Reichel, M.; Currie, P.J. (2014). "Multivariate analysis of isolated tyrannosaurid teeth from the Danek Bonebed, Horseshoe Canyon Formation, Alberta, Canada". Canadian Journal of Earth Sciences. 51 (11): 1045–1051. Bibcode:2014CaJES..51.1045T. doi:10.1139/cjes-2014-0072.
- Holtz, Thomas R. (2004). "Tyrannosauroidea". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 111–136. ISBN 978-0-520-24209-8.
- Eberth, D.A. (2005). "The geology." In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 54–82.
- Arbour, V.M.; Burns, M. E.; Sissons, R. L. (2009). "A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus". Journal of Vertebrate Paleontology. 29 (4): 1117–1135. doi:10.1671/039.029.0405. S2CID 85665879.
- Carr, Thomas D. (1999). "Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria)". Journal of Vertebrate Paleontology. 19 (3): 497–520. doi:10.1080/02724634.1999.10011161. S2CID 83744433.
- Lehman, Thomas M.; Carpenter, Kenneth (1990). "A partial skeleton of the tyrannosaurid dinosaur Aublysodon from the Upper Cretaceous of New Mexico". Journal of Paleontology. 64 (6): 1026–1032. doi:10.1017/S0022336000019843. JSTOR 1305741. S2CID 132662000.
- Carr, Thomas D.; Williamson, Thomas E. (2000). "A review of Tyrannosauridae (Dinosauria: Coelurosauria) from New Mexico". In Lucas, Spencer G.; Heckert, Andrew B. (eds.). Dinosaurs of New Mexico. New Mexico Museum of Natural History and Science Bulletin 17. pp. 113–146.
- Carr, T.D.; Williamson, T.E. (2010). "Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea". Journal of Vertebrate Paleontology. 30 (1): 1–16. doi:10.1080/02724630903413032. S2CID 54029279.
- Horner, John R.; Varricchio, David J.; Goodwin, Mark B. (1992). "Marine transgressions and the evolution of Cretaceous dinosaurs". Nature. 358 (6381): 59–61. Bibcode:1992Natur.358...59H. doi:10.1038/358059a0. S2CID 4283438.
- Varricchio, David J. (2001). "Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts". Journal of Paleontology. 75 (2): 401–406. doi:10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2. S2CID 86113170. DOI: 10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2
- Currie, Philip J.; Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; Murphy, Nate (2005). "An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA)". In Carpenter, Kenneth (ed.). The Carnivorous Dinosaurs. Bloomington: Indiana University Press. pp. 313–324. ISBN 978-0-253-34539-4.
- Carr, Thomas D.; Varricchio, David J.; Sedlmayr, Jayc C.; Roberts, Eric M.; Moore, Jason R. (2017). "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system". Scientific Reports. 7: 44942. Bibcode:2017NatSR...744942C. doi:10.1038/srep44942. PMC 5372470. PMID 28358353.
- Stein, Walter W.; Triebold, Michael (2013). "Preliminary Analysis of a Sub-adult Tyrannosaurid Skeleton from the Judith River Formation of Petroleum County, Montana". In J. Michael Parrish; Ralph E. Molnar; Philip J. Currie; Eva B. Koppelhus (eds.). Tyrannosaurid Paleobiology. Bloomington: Indiana University Press. pp. 55–77.
- Larson, P.; Carpenter, K., eds. (2008). Tyrannosaurus rex, the Tyrant King (Life of the Past). Indiana University Press. p. 310. ISBN 9780253350879.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 464pp. ISBN 978-0-671-61946-6.
- Christiansen, Per; Fariña, Richard A. (2004). "Mass prediction in theropod dinosaurs". Historical Biology. 16 (2–4): 85–92. doi:10.1080/08912960412331284313. S2CID 84322349.
- Campione, N. E.; Evans, D. C.; Brown, C. M.; Carrano, M. T. (2014). "Body mass estimation in non‐avian bipeds using a theoretical conversion to quadruped stylopodial proportions". Methods in Ecology and Evolution. 5 (9): 913−923. doi:10.1111/2041-210X.12226. hdl:10088/25281.
- "Candeiro.vp" (PDF). geology.cz. Retrieved April 11, 2018.
- Molnar, R. E. (2001). "Theropod paleopathology: a literature survey". In Tanke, D. H.; Carpenter, K.; Skrepnick, M. W. (eds.). Mesozoic Vertebrate Life. Bloomington: Indiana University Press. pp. 337–363. ISBN 978-0-253-33907-2.
- Currie, Philip J.; Hurum, Jørn H.; Sabath, Karol (2003). "Skull structure and evolution in tyrannosaurid phylogeny" (PDF). Acta Palaeontologica Polonica. 48 (2): 227–234. Archived from the original (PDF) on October 25, 2007.
- Voris, Jared T.; Therrien, Francois; Zelenitzky, Darla K.; Brown, Caleb M. (2020). "A new tyrannosaurine (Theropoda: Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids". Cretaceous Research. 110: 104388. doi:10.1016/j.cretres.2020.104388. S2CID 213838772.
- Carr, Thomas D.; Williamson, Thomas E.; Schwimmer, David R. (2005). "A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama". Journal of Vertebrate Paleontology. 25 (1): 119–143. doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2. S2CID 86243316.
- Loewen, M.A.; Irmis, R.B.; Sertich, J.J.W.; Currie, P. J.; Sampson, S. D. (2013). Evans, David C (ed.). "Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans". PLoS ONE. 8 (11): e79420. Bibcode:2013PLoSO...879420L. doi:10.1371/journal.pone.0079420. PMC 3819173. PMID 24223179.
- "This Is Our Best Look Yet at a Tyrannosaur's Face". nationalgeographic.com. March 30, 2017. Retrieved April 11, 2018.
- "Scientists discover new dinosaur evolved by anagenesis". upi.com. Retrieved April 11, 2018.
- Tanke, Darren H.; Currie, Philip J. (1998). "Head-biting behavior in theropod dinosaurs: paleopathological evidence" (PDF). Gaia. 15: 167–184. Archived from the original (PDF) on February 27, 2008. [not printed until 2000]
- Eberth, David A.; McCrea, Richard T. (2001). "Were large theropods gregarious?". Journal of Vertebrate Paleontology. 21 (Supplement to Number 3): 46A. doi:10.1080/02724634.2001.10010852. S2CID 220414868.
- Roach, Brian T.; Brinkman, Daniel L. (2007). "A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs". Bulletin of the Peabody Museum of Natural History. 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2. S2CID 84175628. Archived from the original on August 10, 2007.
- Hone, D. W. E.; Tanke, D. H. (2015). "Pre- and postmortem tyrannosaurid bite marks on the remains of Daspletosaurus (Tyrannosaurinae: Theropoda) from Dinosaur Provincial Park, Alberta, Canada". PeerJ. 3: e885. doi:10.7717/peerj.885. PMC 4393819. PMID 25870775.
- Erickson, Gregory M., GM; Makovicky, Peter J.; Currie, Philip J.; Norell, Mark A.; Yerby, Scott A.; Brochu, Christopher A. (2004). "Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs". Nature. 430 (7001): 772–775. Bibcode:2004Natur.430..772E. doi:10.1038/nature02699. PMID 15306807. S2CID 4404887.
- Erickson, Gregory M.; Currie, Philip. J.; Inouye, Brian D.; Wynn, Alice A. (2006). "Tyrannosaur life tables: an example of nonavian dinosaur population biology". Science. 313 (5784): 213–217. Bibcode:2006Sci...313..213E. doi:10.1126/science.1125721. PMID 16840697. S2CID 34191607.
- Wolff, E. D. S.; Salisbury, S. W.; Horner, J. R.; Varricchio, D. J. (2009). "Common Avian Infection Plagued the Tyrant Dinosaurs". PLOS ONE. 4 (9): e7288. Bibcode:2009PLoSO...4.7288W. doi:10.1371/journal.pone.0007288. PMC 2748709. PMID 19789646.
- English, Joseph M.; Johnston, Stephen T. (2004). "The Laramide Orogeny: what were the driving forces?" (PDF). International Geology Review. 46 (9): 833–838. Bibcode:2004IGRv...46..833E. doi:10.2747/0020-6814.46.9.833. S2CID 129901811. Archived from the original (PDF) on June 7, 2011. Retrieved March 13, 2022.
- Eberth, David A. (1997). "Judith River Wedge". In Currie, Philip J.; Padian, Kevin (eds.). Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 199–204. ISBN 978-0-12-226810-6.
- Rogers, Raymond R. (1997). "Two Medicine Formation". In Currie, Philip J.; Padian, Kevin (eds.). Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 199–204. ISBN 978-0-12-226810-6.
- Rogers, Raymond R. (1990). "Taphonomy of three dinosaur bonebeds in the Upper Cretaceous Two Medicine Formation of northwestern Montana: evidence for drought-induced mortality". PALAIOS. 5 (5): 394–413. doi:10.2307/3514834. JSTOR 3514834.
- Falcon-Lang, Howard J. (2003). "Growth interruptions in silicified conifer woods from the Upper Cretaceous Two Medicine Formation, Montana, USA: implications for palaeoclimate and dinosaur palaeoecology". Palaeogeography, Palaeoclimatology, Palaeoecology. 199 (3–4): 299–314. Bibcode:2003PPP...199..299F. doi:10.1016/S0031-0182(03)00539-X.
- Farlow, James O.; Pianka, Eric R. (2002). "Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs". Historical Biology. 16 (1): 21–40. doi:10.1080/0891296031000154687. S2CID 18114585.
- Snively, Eric; Henderson, Donald M.; Phillips, Doug S. (2006). "Fused and vaulted nasals of tyrannosaurid dinosaurs: implications for cranial strength and feeding mechanics" (PDF). Acta Palaeontologica Polonica. 51 (3): 435–454. Archived from the original (PDF) on July 4, 2007.
- Farlow, James O. (1976). "Speculations about the diet and foraging behavior of large carnivorous dinosaurs". American Midland Naturalist. 95 (1): 186–191. doi:10.2307/2424244. JSTOR 2424244.
- Jacobsen, A. R. (2001). "Tooth-marked small theropod bone: An extremely rare trace". In Tanke, D. H.; Carpenter, K.; Skrepnick, M. W. (eds.). Mesozoic Vertebrate Life. Bloomington: Indiana University Press. pp. 58–63. ISBN 978-0-253-33907-2.
External links
- Discussion and specimen list at The Theropod Database.
- Skull image of the Dinosaur Park Formation Daspletosaurus at The Graveyard.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||













