Pika
A pika (/ˈpaɪkə/ PY-kə or /ˈpiːkə/ PEE-kə; archaically spelled pica) is a small, mountain-dwelling mammal found in Asia and North America. With short limbs, very round body, an even coat of fur, and no external tail, they resemble their close relative, the rabbit, but with short, rounded ears.[3] The large-eared pika of the Himalayas and nearby mountains is found at heights of more than 6,000 m (20,000 ft), among the highest of any mammal.
| Pika[1] | |
|---|---|
_with_a_mouthful_of_flowers.jpg.webp) | |
| American pika (Ochotona princeps) | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Lagomorpha |
| Family: | Ochotonidae Thomas, 1897 |
| Genus: | Ochotona Link, 1795 |
| Type species | |
| Ochotona daurica Link, 1795 (Lepus dauuricus Pallas, 1776) | |
| Species | |
|
See text | |
Pikas prefer rocky slopes and graze on a range of plants, mostly grasses, flowers and young stems. In the autumn they pull hay, soft twigs and other stores of food into their burrows to eat during the long, cold winter.[4] The pika is also known as the whistling hare because of its high-pitched alarm call when diving into its burrow.
The name ‘pika’ appears to be derived from the Tungus piika,[5] and the scientific name Ochotona is from the Mongolian word ogutun-a, оготно, which means pika.[6] It is used for any member of the Ochotonidae,[7] a family within the order of lagomorphs which also includes the Leporidae (rabbits and hares). Only one genus, Ochotona,[7] is extant within the family, covering 37 species, though many fossil genera are known. Another species, the Sardinian pika, belonging to the separate genus Prolagus, has become extinct within the last 2000 years owing to human activity.
The two species found in North America are the American pika, found primarily in the mountains of the western United States and far southwestern Canada, and the collared pika of northern British Columbia, the Yukon, western Northwest Territories and Alaska.
Habitat

Pikas are native to cold climates in Asia and North America. Most species live on rocky mountainsides, where numerous crevices are available for their shelter, although some pikas also construct crude burrows. A few burrowing species are native to open steppe land. In the mountains of Eurasia pikas often share their burrows with snowfinches, which build their nests there.[8] Pikas require cold temperatures to live, and can die if exposed to temperatures above 25.5 °C (77.9 °F).[9] Changing temperatures have forced some pika populations to restrict their ranges to even higher elevations.[10]
Characteristics

Pikas are small mammals, with short limbs and rounded ears. They are about 15 to 23 cm (6 to 9 in) in body length and weigh between 120 and 350 g (4 and 12+1⁄2 oz), depending on species. Like rabbits, after eating they initially produce soft green feces, which they eat again to take in further nutrition before producing the final solid fecal pellets. Collared pikas have been known to store dead birds in their burrows for food during winter and eat the feces of other animals.[11]
These animals are herbivores and feed on a wide variety of plant matter, including forbs, grasses, sedges, shrub twigs, moss and lichens. As with other lagomorphs, pikas have gnawing incisors and no canines, although they have fewer molars than rabbits; they have a dental formula of: 2.0.3.21.0.2.3. Another similarity that pikas share with other lagomorphs is that the bottom of their paws are covered with fur and therefore lack paw pads.[12]
Rock-dwelling pikas have small litters of fewer than five young, whilst the burrowing species tend to give birth to more young and to breed more frequently, possibly owing to a greater availability of resources in their native habitats. The young are born after a gestation period of between 25 and 30 days.[8]
Activity


Pikas are active during daylight (diurnal) or twilight hours (crepuscular), with higher-elevation species generally being more active during the daytime. They show their peak activity just before the winter season. Pikas do not hibernate so they generally spend time during the summer collecting and storing food for the winter. Each rock-dwelling pika stores its own ‘haypile’ of dried vegetation, whilst burrowing species often share food stores with their burrow mates. Haying behavior is more prominent at higher elevations. Many of the vocalizations and social behaviors that pikas exhibit are related to haypile defense.
The impact of human activity on the tundra ecosystems where pikas live has been recorded dating back to the 1970s.[13] Rather than hibernate during winter, pikas forage for grasses and other forms of plant matter and stash these findings in protected dens in a process called 'haying' which they sustain themselves with during winter seasons.[14] Sometimes pikas think that humans are predators, so they sometimes respond to humans as they do to other species that may actually be preying on the pikas. Such interactions with humans have been linked to pikas having reduced amounts of foraging time, consequentially limiting the amount of food they can stockpile for winter months.[15] Pikas are unable to withstand surrounding temperatures above 25 °C (77 °F) so they must spend their time in shaded regions and out of direct sunlight when temperatures are high.[15] A link has also been found between temperature increases and lost foraging time, where for every increase of 1°C to the ambient temperature in alpine landscapes home to pikas those pikas lose 3% of their foraging time.[15]
Eurasian pikas commonly live in family groups and share duties of gathering food and keeping watch. Some species are territorial. North American pikas (O. princeps and O. collaris) are asocial, leading solitary lives outside the breeding season.[16]
Dialects
Pikas have distinct calls, which vary in duration. The call can be short and quick, a little longer and more drawn out or long songs. The short calls are an example of geographic variation. The pikas determine the appropriate time to make short calls by listening for cues for sound localization.[17] The calls are used for individual recognition, predator warning signals, territory defense, or as a way to attract the opposite sex.[18] There are also different calls depending on the season. In the spring the songs become more frequent during the breeding season. In late summer the vocalizations become short calls. Through various studies, the acoustic characteristics of the vocalizations can be a useful taxonomic tool.[19]
Lifespan
The average lifespan of pikas in the wild is roughly seven years. A pika's age may be determined by the number of adhesion lines on the periosteal bone on the lower jaw. The lifespan does not differ between the sexes.[20]
Species
The 34 species currently recognized are:
- Order Lagomorpha[1]
- Family Ochotonidae: pikas
- Genus Ochotona
- Subgenus Conothoa: mountain pikas
- Chinese red pika, O. erythrotis
- Forrest's pika, O. forresti
- Gaoligong pika (O. gaoligongensis) and black pika (O. nigritia) are now thought to be conspecific with O. forresti
- Glover's pika, O. gloveri
- Muli pika (O. muliensis) is now thought to be conspecific with O. gloveri
- Ili pika, O. iliensis
- Koslov's pika, O. koslowi
- Ladak pika, O. ladacensis
- Large-eared pika, O. macrotis
- Royle's pika, O. roylei
- Himalayan pika (O. himalayana) is now thought to be conspecific with O. roylei
- Turkestan red pika, O. rutila
- Subgenus Alienauroa
- Yellow pika, O. huanglongensis
- Sacred pika, O. sacraria
- Flat-headed pika, O. flatcalvariam
- Subgenus Ochotona: shrub-steppe pikas
- Gansu pika or gray pika, O. cansus
- Plateau pika or black-lipped pika, O. curzoniae
- Daurian pika, O. dauurica
- Nubra pika, O. nubrica
- Steppe pika, O. pusilla
- Qionglai pika, O. qionglaiensis
- Afghan pika, O. rufescens
- Sijin pika, O. sikimaria
- Tsing-ling pika, O. syrinx
- Moupin pika, O. thibetana
- Thomas's pika, O. thomasi
- Subgenus Pika: northern pikas
- Alpine pika or Altai pika, O. alpina
- Helan Shan pika or silver pika, O. argentata
- Collared pika, O. collaris
- Korean pika, O. coreana
- Hoffmann's pika, O. hoffmanni
- Northern pika or Siberian pika, O. hyperborea
- Manchurian pika, O. mantchurica
- Kazakh pika, O. opaca
- Pallas's pika, O. pallasii
- American pika, O. princeps
- Turuchan pika, O. turuchanensis
- Subgenus Conothoa: mountain pikas
- Genus Ochotona
- Family Ochotonidae: pikas
Extinct species
Many fossil forms of Ochotona are described in the literature, from the Miocene epoch to the early Holocene (extinct species) and present (16.4-0 Ma[2]). They lived in Europe, Asia, and North America.Some species listed below are common for Eurasia and North America (O. gromovi, O. tologoica, O. zazhigini, and probably O. whartoni).
- Eurasia
- large forms
- †Ochotona chowmincheni (China: Baode area, late Miocene)[2][21][22]
- †Ochotona gromovi (Asia, Pliocene, see also North America)[22]
- †Ochotona gudrunae (China: Shanxi, early Pleistocene)[2][21][22]
- †Ochotona guizhongensis (Tibet, late Miocene)[2][22][23]
- †Ochotona lagreli (China: Inner Mongolia, late Miocene to late Pliocene)[2][21][22][23]
- †Ochotona magna (China, early Pleistocene)[2][22][24]
- †Ochotona tologoica (Transbaikalia, Pliocene, see also North America)[2][22][24]
- †Ochotona transcaucasica (Transcaucasia: eastern Georgia and Azerbaijan, Transbaikal and probably southern Europe, early to late Pleistocene)[2][21][22]
- †Ochotona ursui (Romania, Pliocene)[2][22]
- †Ochotona zasuchini (Transbaikalia, Pleistocene)[2][22][24]
- †Ochotona zazhigini (Asia, Pliocene, see also North America)[2][22]
- †Ochotona zhangi (China, Pleistocene)[2][22][24]
- medium-sized forms
- †Ochotona agadjianiani (Asia, Pliocene)[22]
- †Ochotona antiqua (Moldavia, Ukraine, and the Russian Plain, Caucasus, and probably Rhodes, late Miocene to Pliocene)[2][21][22]
- †Ochotona azerica (Transcaucasia: Azerbaijan,[25] middle Pliocene)[2][24]
- †Ochotona lingtaica (Asia, Pliocene)[2][22]
- †Ochotona dodogolica (Asia: western Transbaikalia, Pleistocene)[21][22]
- †Ochotona nihewanica (China: Hebei, early Pleistocene)[2][21][22][26]
- †Ochotona plicodenta (Asia, Pliocene)[2][22]
- †Ochotona polonica (Europe: Poland, Germany, France, Pliocene)[2][21][22]
- small-sized forms
- †Ochotona bazarovi (Asia, upper Pliocene)[2][24][26]
- †Ochotona dehmi (Germany: Schernfeld, Pleistocene)[2][22]
- †Ochotona filippovi (Siberia, Pleistocene)[2][26]
- †Ochotona gracilis (Asia, Pliocene)[2][22]
- †Ochotona horaceki (Slovakia: Honce, Pleistocene)[2][22]
- †Ochotona minor (China, late Miocene)[2][22][23]
- †Ochotona sibirica (Asia, Pliocene)[2][22]
- †Ochotona valerotae (France: Valerots site, Pleistocene)[2][22]
- †Ochotona youngi (Asia, Pliocene)[22]
and others.[2][22]
- other examples
- †Ochotona agadzhaniani (Transcaucasia: Armenia, Pliocene)[2]
- †Ochotona alaica (Asia: Kyrgyzstan, Pleistocene)[2]
- †Ochotona (Proochotona) eximia (Moldova, Ukraine, Russia, Kazakhstan, Miocene to Pliocene)[2]
- †Ochotona (Proochotona) gigas (Ukraine, Pliocene)[2]
- †Ochotona gureevi (Transbaikalia, middle Pliocene)[2][24]
- †Ochotona hengduanshanensis (China, Pleistocene)[2]
- †Ochotona intermedia (Asia, Pliocene)[2][22]
- †Ochotona (Proochotona) kalfaense (Europe: Moldova, Miocene)[2]
- †Ochotona (Proochotona) kirgisica (Asia: Kyrgyzstan, Pliocene)[2]
- †Ochotona kormosi (Hungary, Pleistocene)[2][21]
- †Ochotona (Proochotona) kurdjukovi (Asia: Kyrgyzstan, Pliocene)[2]
- †Ochotona largerli (Georgia, Pleistocene)[2]
- †Ochotona lazari (Ukraine, Pleistocene)[2]
- †Ochotona mediterranensis (Turkey, Pliocene)[2]
- †Ochotona ozansoyi (Turkey, Miocene)[2]
- †Ochotona pseudopusilla (Ukraine and Russian Plain, Pleistocene)[2][22]
- †Ochotona spelaeus (Ukraine, late Pleistocene)[2][27]
- †Ochotona tedfordi (China: Yushe Basin, late Miocene)[2][21]
- †Ochotona cf. whartoni (Irkutsk Oblast and Yakutia, Pleistocene, see also North America)[22]
- †Ochotona zabiensis (southern Poland, early Pleistocene)[2][21]
- †Ochotona sp. (Greece: Maritsa, Pliocene)[22]
- †Ochotona sp. (Hungary: Ostramos, Pleistocene)[22]
- †Ochotona sp. (Siberia, Pleistocene)[22]
- †Ochotona sp. (Yakutia, Pleistocene)[22]
- large forms
- North America
- †Ochotona gromovi (US: Colorado, Pliocene, see also Eurasia)[2]
- †Ochotona spanglei (US, late Miocene or early Pliocene)[2][22][26][28][29][n 1]
- †Ochotona tologoica (US: Colorado, Pliocene, see also Eurasia)[2]
- †Ochotona whartoni (giant pika, US, Canada, Pleistocene to early Holocene, see also Eurasia)[2][22][26][30][31][n 2]
- †Ochotona wheatleyi (US: Alaska, Pliocene, late Pleistocene)[2]
- †Ochotona zazhigini (US: Colorado, Pleistocene, see also Eurasia)[2]
- extinct small pikas similar to the O. pusilla group (Pleistocene)[22][26]
Paleontologists have also described multiple forms of pika not referred to specific species (Ochotona indet.) or not certainly identified (O. cf. antiqua, O. cf. cansus, O. cf. daurica, O. cf. eximia, O. cf. gromovi, O. cf. intermedia, O. cf. koslowi, O. cf. lagrelii, O. cf. nihewanica). The statuses of Ochotona (Proochotona) kirgisica and O. spelaeus are uncertain.[2]
The "pusilla" group of pikas is characterized by archaic (plesiomorphic) cheek teeth and small size.[26]
The North American species migrated from Eurasia. They invaded the New World twice:
- O. spanglei during the latest Miocene or early Pliocene, followed by a roughly three-million-year-long gap in the known North American pikas record[22]
- O. whartoni (giant pika) and small pikas via the Bering Land Bridge during the earliest Pleistocene[22]
Ochotona cf. whartoni and small pikas of the O. pusilla group are also known from Siberia. The extant, endemic North American species appeared in the Pleistocene. The North American collared pika (O. collaris) and American pika (O. princeps) have been suggested to have descended from the same ancestor as the steppe pika (O. pusilla).[22]
The range of Ochotona was larger in the past, with both extinct and extant species inhabiting Western Europe and Eastern North America, areas that are currently free of pikas. Pleistocene fossils of the extant steppe pika O. pusilla currently native to Asia have been found also in many countries of Europe from the United Kingdom to Russia and from Italy to Poland, and the Asiatic extant northern pika O. hyperborea in one location in the middle Pleistocene United States.[2]
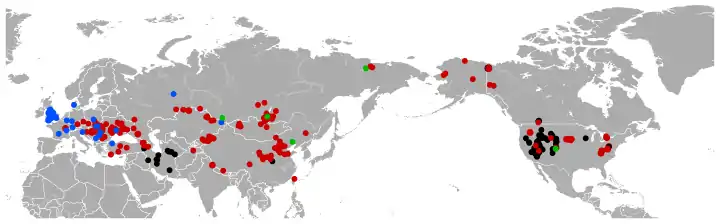

Other genera of ochotonids (currently living only Ochotonidae) include except Ochotona (pika) extinct †Albertona, †Alloptox, †Amphilagus, †Australagomys, †Austrolagomys, †Bellatona, †Bellatonoides, †Bohlinotona, †Cuyamalagus, †Desmatolagus, †Eurolagus, †Gripholagomys, †Gymnesicolagus, †Hesperolagomys, †Heterolagus, †Kenyalagomys, †Lagopsis, †Marcuinomys, †Ochotonoides, †Ochotonoma, †Oklahomalagus, †Oreolagus, †Paludotona, †Piezodus, †Plicalagus, †Pliolagomys, †Prolagus, †Proochotona (syn. Ochotona), †Pseudobellatona, †Ptychoprolagus, †Russellagus, †Sinolagomys and †Titanomys.[2] The earliest one is Desmatolagus (middle Eocene to Miocene, 42.5–14.8 Ma[2]), usually included in the Ochotonidae, sometimes in Leporidae or in neither ochotonid nor leporid stem-lagomorphs.[33]
Ochotonids appeared in Asia between the late Eocene and the early Oligocene, and continued to develop along with increased distribution of C3 grasses in previously forest dominated areas under the "climatic optimum" from the late Oligocene to middle Miocene. They thrived in Eurasia, North America, and even Africa. The peak of their diversity occurred during the period from the early Miocene to middle Miocene, most of them became extinct during the transition from the Miocene to Pliocene, what was accompanied by diversity increase in the leporids. It has been proposed that this switch between ochotonids and larger leporids was caused by expansion of C4 plants (particularly the Poaceae) related to global cooling in the late Miocene, since extant pikas reveal a strong preference for C3 plants (Asteraceae, Rosaceae, and Fabaceae, many of them C3). Replacement of large areas of forests by open grassland first started probably in North America and is called sometimes "nature's green revolution".[2]
Notes
- Ochotona spanglei in the Paleobiology Database.[29][pdb 1][pdb 2][pdb 3]
- Ochotona whartoni in the Paleobiology Database.[31][pdb 4][pdb 5][pdb 6][pdb 7][pdb 8][pdb 9][pdb 10]
- The coordinates of additional fossils not listed in the xls file attached to Ge and all paper[2] were taken from the Paleobiology Database.[32][pdb 11][pdb 12][pdb 13][pdb 14][pdb 15][pdb 16][pdb 17][pdb 18][pdb 19][pdb 20][pdb 6][pdb 7][pdb 21][pdb 22][pdb 5][pdb 23][pdb 24][pdb 25][pdb 26][pdb 27][pdb 28][pdb 29][pdb 30]
References
- Hoffman, R.S.; Smith, A.T. (2005). "Order Lagomorpha". In Wilson, D.E.; Reeder, D.M (eds.). Mammal Species of the World: A Taxonomic and Geographic Reference (3rd ed.). Johns Hopkins University Press. pp. 185–193. ISBN 978-0-8018-8221-0. OCLC 62265494.
- Ge, Deyan; Wen, Zhixin; Xia, Lin; Zhang, Zhaoqun; Erbajeva, Margarita; Huang, Chengming; Yang, Qisen (April 3, 2013). "Evolutionary History of Lagomorphs in Response to Global Environmental Change". PLOS ONE. 8 (4:e59668): e59668. Bibcode:2013PLoSO...859668G. doi:10.1371/journal.pone.0059668. PMC 3616043. PMID 23573205. Table_S1.xls Archived 2014-05-22 at the Wayback Machine
- Melissa Breyer (2 September 2016). "Meet the 'mouse-bunny' that could vanish from the US". treehugger.
- Walters, Martin (2005). Encyclopedia of animals. Parragon. p. 203. ISBN 978-1-40545-669-2.
- Harper, Douglas. "pika". Online Etymology Dictionary.
- General pika information Archived 2017-05-10 at the Wayback Machine. twycrosszoo.org
- Lydekker, Richard (1911). . In Chisholm, Hugh (ed.). Encyclopædia Britannica. Vol. 21 (11th ed.). Cambridge University Press. p. 575.
- Kawamichi, Takeo (1984). Macdonald, D. (ed.). The Encyclopedia of Mammals. New York: Facts on File. pp. 726–727. ISBN 978-0-87196-871-5.
- "American Pika". U.S. Fish and Wildlife Service. Retrieved 2018-11-18.
- Erb, Liesl P; Ray, Chris; Guralnick, Robert (2011-09-01). "On the generality of a climate-mediated shift in the distribution of the American pika (Ochotona princeps)". Ecology. 92 (9): 1730–1735. doi:10.1890/11-0175.1. PMID 21939069.
- Leininger, Charlene (2009) Ochotona collaris Archived 2013-06-28 at the Wayback Machine. Animal Diversity Web
- "Pika | mammal". Encyclopedia Britannica. Retrieved 2021-06-26.
- Brown, R. W., R. S. Johnston, and K. Van Cleve. "Rehabilitation problems of Arctic and alpine regions." Reclamation of drastically disturbed lands (1978): 23-44.
- Dearing, M. Denise. "The function of haypiles of pikas (Ochotona princeps)." Journal of Mammalogy 78.4 (1997): 1156-1163. APA
- Stafl, Natalie; O'Connor, Mary I. (2015-08-01). "American Pikas' (Ochotona princeps) Foraging Response to Hikers and Sensitivity to Heat in an Alpine Environment". Arctic, Antarctic, and Alpine Research. 47 (3): 519–527. doi:10.1657/AAAR0014-057. ISSN 1523-0430. S2CID 86263545.
- Leininger, Charlene (2009). "Ochotona collaris". Animal Diversity Web. Retrieved March 23, 2021.
- Conner, Douglas A. (1982-02-25). "Geographic Variation in Short Calls of Pikas (Ochotona princeps)". Journal of Mammalogy. 63 (1): 48–52. doi:10.2307/1380670. JSTOR 1380670.
- Trefry, Sarah A.; Hik, David S. (2009). "Variation in pika (Ochotona collaris, O. princeps) vocalizations within and between populations". Ecography. 33 (4): 784–795. doi:10.1111/j.1600-0587.2009.05589.x.
- Somers, Preston (1973). "Dialects in southern Rocky Mountain pikas, Ochotona princeps (Lagomorpha)". Animal Behaviour. 21: 124–137. doi:10.1016/S0003-3472(73)80050-8.
- Barker, Jennifer M; Boonstra, Rudy; Schulte-Hostedde, Albrecht I (2003-10-01). "Age determination in yellow-pine chipmunks (Tamias amoenus): a comparison of eye lens masses and bone sections". Canadian Journal of Zoology. 81 (10): 1774–1779. doi:10.1139/z03-173.
- Fostowicz-Frelik, Łucja; Frelik, Grzegorz; Gasparik, Mihály (October 2010). "Morphological phylogeny of pikas (Lagomorpha: Ochotona), with a description of a new species from the Pliocene/Pleistocene transition of Hungary". Proceedings of the Academy of Natural Sciences of Philadelphia. 159: 97–117. doi:10.1635/053.159.0107. JSTOR 41446115. S2CID 83700561.
- Erbajeva, Margarita A.; Mead, Jim I.; Alexeeva, Nadezhda V.; Angelone, Chiara; Swift, Sandra L. (2011). "Taxonomic diversity of Late Cenozoic Asian and North American ochotonids (an overview)" (PDF). Palaeontologia Electronica: 1–9. Archived (PDF) from the original on April 14, 2014. Retrieved April 13, 2014.
- Cai, Baoquan (1989). "Fossil Lagomorpha from the Late Pliocene of Yangyuan and Yuxian counties, Hebei Province" (PDF). Vertebrata PalAsiatica. XXVII (3): 170–181. Archived (PDF) from the original on March 5, 2016. Retrieved May 20, 2014.
Translated by Will Downs Department of Geology Bilby Research Center Northern Arizona University October, 1990
- Erbajeva, Margarita A.; Zheng, Shaohua (30 June 2005). "New data on Late Miocene – Pleistocene ochotonids (Ochotonidae, Lagomorpha) from North China" (PDF). Acta Zoologica Cracoviensia. 48A (1–2): 93–117. doi:10.3409/173491505783995734. Archived (PDF) from the original on May 10, 2017. Retrieved May 20, 2014.
- Čermák, Stanislav; Obuch, Ján; Benda, Petr (2006). "Notes on the genus Ochotona in the Middle East (Lagomorpha: Ochotonidae)" (PDF). Lynx. 37: 51–66. ISSN 0024-7774. Archived from the original (PDF) on May 22, 2014. Retrieved May 22, 2014.
- Erbajeva, Margarita A.; Mead, Jim I.; Swift, Sandra L. (2003). "Evolution and development of Asian and North American ochotonids" (PDF). Occasional Papers in Earth Sciences No. 5: 33–34. Archived from the original (PDF) on March 31, 2014. Retrieved April 13, 2014.
3rd INTERNATIONAL MAMMOTH CONFERENCE, 2003: PROGRAM AND ABSTRACTS, Edited by John E. Storer
- Rekovets, Leonid (2003). "Mammoth (Mammuthus primigenius) in the periglacial faunas of Ukraine" (PDF). Occasional Papers in Earth Sciences No. 5: 130–131. Archived from the original (PDF) on March 31, 2014. Retrieved April 13, 2014.
3rd INTERNATIONAL MAMMOTH CONFERENCE, 2003: PROGRAM AND ABSTRACTS, Edited by John E. Storer
- Shotwell, J. Arnold (1956). "Hemphillian mammalian assemblage from northeastern Oregon". Geological Society of America Bulletin. 67 (6): 717–738. Bibcode:1956GSAB...67..717S. doi:10.1130/0016-7606(1956)67[717:HMAFNO]2.0.CO;2.
- "Ochotona spanglei Shotwell 1956". The Paleobiology Database. Archived from the original on April 15, 2014.
- Guthrie, R.D.; Matthews, John V. Jr. (1971). "The Cape Deceit fauna—Early pleistocene mammalian assemblage from the Alaskan arctic". Quaternary Research. 1 (4): 474–510. Bibcode:1971QuRes...1..474G. doi:10.1016/0033-5894(71)90060-3. S2CID 86601856.
- "Ochotona whartoni Guthrie and Matthews, Jr. 1971 (pika)". The Paleobiology Database. Archived from the original on April 14, 2014.
- "Ochotona Link 1795 (pika)". The Paleobiology Database.
- Hordijk, Kees (2010). Perseverance of pikas in the Miocene : interplay of climate and competition in the evolution of Spanish Ochotonidae (Lagomorpha, Mammalia). Geologica Ultraiectina. Vol. 333. Departement Aardwetenschappen. hdl:1874/197550. ISBN 978-90-5744-194-3.
document type Dissertation full text
{{cite book}}: External link in|quote=
Additional references of the Paleobiology Database
- Shotwell, J. A. (1956). "Hemphillian mammalian assemblage from Northeastern Oregon". Geological Society of America Bulletin. 67 (6): 717. Bibcode:1956GSAB...67..717S. doi:10.1130/0016-7606(1956)67[717:hmafno]2.0.co;2. [J. Alroy/J. Alroy]
- Voorhies, M. R. (1990). Gustavson, T. C. (ed.). Bureau of Economic Geology Guidebook. [J. Alroy/J. Alroy]
- Additional contributors to utilized records of Paleobiology Database (authorizers supplying these records) include John Alroy.
- Guthrie, R. D.; Matthews, J. V. Jr. (1971). "The Cape Deceit fauna—Early pleistocene mammalian assemblage from the Alaskan arctic". Quaternary Research. 1 (4): 474–510. Bibcode:1971QuRes...1..474G. doi:10.1016/0033-5894(71)90060-3. S2CID 86601856. [J. Alroy/J. Alroy]
- Jopling, A. V.; et al. (1981). "Stratigraphic, Sedimentological and Faunal Evidence for the Occurrence of Pre-Sangamonian Artefacts in Northern Yukon". Arctic. 34 (1). doi:10.14430/arctic2499. [J. Alroy/J. Alroy]
- Harington, C. R. (1978). "Quaternary vertebrate faunas of Canada and Alaska and their suggested chronological sequence". Syllogeus. 15. [J. Alroy/J. Alroy]
- Harington, C. R. (1990). "Vertebrates of the last interglaciation in Canada: A review" (PDF). Geographie Physique et Quaternaire. 44 (3): 375. doi:10.7202/032837ar. [J. Alroy/J. Alroy/M. Uhen]
- Storer, J. E. (2004). "A Middle Pleistocene (late Irvingtonian) mammalian fauna from Thistle Creek, Klondike Goldfields region of Yukon Territory, Canada". Paludicola. 4 (4): 137–150. [J. Alroy/J. Alroy]
- Tedford, R. H.; Wang, X; Taylor, B. E. (2009). "Phylogenetic Systematics of the North American Fossil Caninae (Carnivora: Canidae)". Bulletin of the American Museum of Natural History. 325: 1–218. doi:10.1206/574.1. hdl:2246/5999. S2CID 83594819. [J. Marcot/J. Marcot]
- Additional contributors to utilized records of Paleobiology Database (authorizers supplying these records) include John Alroy, Jonathan Marcot.
- Barnosky, A. D.; Rasmussen, D. L. (1988). "Middle Pleistocene arvicoline rodents and environmental change at 2900-meters elevation, Porcupine Cave, South Park, Colorado". Annals of Carnegie Museum. 57 (12): 267–292. doi:10.5962/p.330577. [J. Alroy/J. Alroy]
- Belyaeva, E. I. (1948). Catalogue of Tertiary Fossil Sites of the Land Mammals in the U.S.S.R. [M. Uhen/M. Uhen]
- Bonifay, M. F. (1973). "Principaux gisements paléontologiques Français du Pléistocene Moyen: Essai de classification". Le Quaternaire: 41–50. [A. Turner/H. O'Regan/H. O'Regan]
- Cai, B. (1987). "A preliminary report on the Late Pliocene Micromammalian fauna from Yangyuan and Yuxian, Hebei". Vertebrata PalAsiatica. 25 (2): 124–136. [A. Turner/H. O'Regan/H. O'Regan]
- Deng, T.; Wang, X.; Fortelius, M.; Li, Q.; Wang, Y.; Tseng, Z. J.; Takeuchi, G. T.; Saylor, J. E.; Säilä, L. K.; Xie, G. (2011). "Out of Tibet: Pliocene woolly rhino suggests high-plateau origin of Ice Age megaherbivores". Science. 333 (6047): 1285–1288. Bibcode:2011Sci...333.1285D. doi:10.1126/science.1206594. PMID 21885780. S2CID 8913866. [J. Alroy/J. Alroy/J. Alroy]
- Erbaeva, M. A. (1986). "The Late Cenozoic Faunistic complexes of Transbaikalia with special reference to the micromammalia". Quatärpaläontologie. 6: 25–28. [A. Turner/H. O'Regan/H. O'Regan]
- Frazier, M. K. (1977). "New Records of Neofiber leonardi (Rodentia: Cricetidae) and the Paleoecology of the Genus". Journal of Mammalogy. 58 (3): 368–373. doi:10.2307/1379335. JSTOR 1379335. [M. Uhen/M. Shalap]
- Gidley, J. W. (1913). "Preliminary report on a recently discovered Pleistocene cave deposit near Cumberland, Maryland". Proceedings of the United States National Museum. 46 (2014): 93–102. doi:10.5479/si.00963801.46-2014.93. hdl:2027/hvd.32044107347718. [M. Uhen/M. Shalap/M. Shalap]
- Grady, F.; Garton, E. R. (2000). "Paleontology and historic field trip of the John Guilday Cave Preserve (Trout Rock)". Bulletin – West Virginia Speleological Survey. 14: 241–244. [M. Uhen/M. Shalap/M. Shalap]
- Guilday, J. E. (1979). "Eastern North American Pleistocene Ochotona (Lagomorpha: Mammalia). Carnegie Museum of Natural History". Annals of Carnegie Museum. 48 (24). doi:10.5962/p.330836. S2CID 251525193. [J. Alroy/J. Alroy]
- Janossy, D. (1970). "Ein neuer Eomyide (Rodentia, Mammalia) aus dem Ältestpleistozän ("Oberes Villafrankium", Villanyium) des Osztramos (Nordostungarn); (A new Eomyid (Rodentia, Mammalia) from the lowermost Pleistocene (upper Villafranchian) from Osztramos mountain (Northeastern Hungary)". Annales Historico-Naturales Musei Nationalis Hungarici. 62: 99–113. [J. Alroy/S. Kuemmell/S. Kuemmell]
- Janossy, D. (1986). Pleistocene vertebrate faunas of Hungary. Developments in Palaeontology and Stratigraphy. Vol. 8. Amsterdam: Elsevier. ISBN 978-0-444-99526-1. [A. Turner/H. O'Regan/H. O'Regan]
- Kurten, B.; Anderson, E. (1980). Pleistocene mammals of North America. ISBN 978-0231037334. [J. Alroy/J. Alroy/J. Alroy]
- Mead, J. I.; Grady, F. (1996). "Ochotona (Lagomorpha) from late Quaternary cave deposits in eastern North America". Quaternary Research. 45 (1): 93–101. Bibcode:1996QuRes..45...93M. doi:10.1006/qres.1996.0009. S2CID 128811270. [J. Alroy/J. Alroy/J. Alroy]
- Qiu, Z. (1987). "Neogene micromammals of China". Whyte, P., ed. Paleoenvironment of East Asia from the Mid-Tertiary, Second International Conference on the Paleoenvironment of East Asia. 77 (1–2): 834–848. [W. Clyde/J. Finarelli/W. Clyde]
- Rasmussen, D. L. (1974). "New Quaternary mammal localities in the upper Clark Fork River valley, western Montana". Northwest Geology. 3: 62–70. [M. Uhen/C. Peredo]
- Sotnikova, M.V.; Dodonov, A.E.; Pen'kov, A.V. (1997). "Upper Cenozoic bio-magnetic stratigraphy of Central Asian mammalian localities". Palaeogeography, Palaeoclimatology, Palaeoecology. 133 (3–4): 243–258. Bibcode:1997PPP...133..243S. doi:10.1016/s0031-0182(97)00078-3. [A. Turner/H. O'Regan/H. O'Regan]
- Terzea, E. (1996). "Biochronology of the Pleistocene deposits at Betfia (Bihor, Romania)". Acta Zoologica Cracovensia. 39 (1): 531–540. [A. Behrensmeyer/H. O'Regan/H. O'Regan]
- Winkler, A. J.; Grady, F. (1990). "The middle Pleistocene rodent Atopomys (Cricetidae: Arvicolinae) from the eastern and south-central United States". Journal of Vertebrate Paleontology. 10 (4): 484–490. doi:10.1080/02724634.1990.10011831. [J. Alroy/J. Alroy]
- Additional contributors to utilized records of Paleobiology Database (authorizers supplying these records) include John Alroy, Anna Behrensmeyer, Will Clyde, Alan Turner, Mark Uhen.
Further reading
- Orr, Robert Thomas (1977). The Little-known Pika (illustrated ed.). New York: Macmillan. ISBN 9780025939608.
External links
- The trek of the pika, by Michael Morris, Parks Canada, Mount Revelstoke and Glacier National Parks. (includes sound file)