Saurolophus
Saurolophus (/sɔːˈrɒləfəs/; meaning "lizard crest") is a genus of large hadrosaurid dinosaur from the Late Cretaceous period of Asia and North America, that lived in what is now the Horseshoe Canyon and Nemegt formations about 70 million to 68 million years ago. It is one of the few genera of dinosaurs known from multiple continents. The type species, S. osborni, was described by Barnum Brown in 1912 from Canadian fossils. A second valid species, S. angustirostris, is represented by numerous specimens from Mongolia, and was described by Anatoly Konstantinovich Rozhdestvensky.
| Saurolophus Temporal range: Late Cretaceous, ~ | |
|---|---|
 | |
| Mounted S. angustirostris skeleton at Nagoya City Science Museum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | †Ornithischia |
| Clade: | †Ornithopoda |
| Family: | †Hadrosauridae |
| Subfamily: | †Saurolophinae |
| Tribe: | †Saurolophini |
| Genus: | †Saurolophus Brown, 1912 |
| Type species | |
| †Saurolophus osborni Brown, 1912 | |
| Other species | |
| |
Saurolophus is distinguished by a spike-like crest which projects up and back from the skull. It was a herbivorous dinosaur which could move about either bipedally or quadrupedally.
Discovery and history

Barnum Brown recovered the first described remains of Saurolophus in 1911, including a nearly complete skeleton (AMNH 5220). Now on display in the American Museum of Natural History, this skeleton was the first nearly complete dinosaur skeleton from Canada. It was found in rocks of early Maastrichtian age, in the Upper Cretaceous Horseshoe Canyon Formation (then known as the Edmonton Formation) near Tolman Ferry on the Red Deer River in Alberta. Brown wasted little time in describing his material,[1][2] giving it its own subfamily.[3] Saurolophus was an important early reference for other hadrosaurs, as seen in the names of Prosaurolophus ("before Saurolophus") and Parasaurolophus ("near Saurolophus"). However, little additional material has been recovered and described.
Instead, more abundant remains from Asia have provided more data. Initial remains were not promising; a partial fragmentary ischium from Heilongjiang, China, that Riabinin named S. kryschtofovici.[4] Much better remains were soon recovered, though, but from Mongolia's early Maastrichtian-age Nemegt Formation. The 1946–1949 Russian-Mongolian paleontological expeditions recovered the large skeleton that became S. angustirostris as described by Anatoly Rozhdestvensky.[5] Other skeletons from a variety of growth stages have also been discovered, and S. angustirostris is now the most abundant Asian hadrosaurid.[6]
Species

Two species are regarded as valid today: the type species S. osborni, and S. angustirostris. S. osborni (Brown, 1912) is known from a skull and skeleton, two other complete skulls, and skull fragments. S. angustirostris (Rozhdestvensky, 1952) is known from at least 15 specimens.[7] It differs from S. osborni by some details of the skull, as well as in the pattern of scales found in skin impressions. The Mongolian species had a longer skull (by 20%) and the front of the snout (the premaxillary bones) were more upwardly directed.[8] S. angustirostris also had a distinctive row of rectangular scales along the midline of the back and tail, known as 'midline feature-scales'; these are not currently preserved in S. osborni. In S. angustirostris, the scales on the tail flank were arranged in vertical patterns, which may have corresponded to striped coloration in life. This area was covered in radial scale patterns in S. osborni, possibly indicating a more mottled or spotted coloration.[9]

S. kryschtofovici (Riabinin, 1930) is not considered valid; either it is regarded as a dubious name,[10][7] or as a synonym of S. angustirostris[6] (although the name antedates S. angustirostris).[11]
Until a 2011 reevaluation of the species by Phil R. Bell, S. angustirostris was not well-described. No autapomorphies, unique derived traits, had been established distinguishing it from S. osborni. Bell found in a publication earlier in the year that the two previous studies of S. angustirostris, by Rozhdestvensky in 1952, and Maryanska and Osmolska in 1981, do not provide a comprehensive enough description to compare the species with S. osborni.[8]
In 1939–40, two partial skeletons were found in the late Maastrichtian age Moreno Formation of California. These specimens were referred to cf. Saurolophus sp. In 2010, one of the skulls was instead assigned to Edmontosaurus.[12] A 2013 study placed the two specimens in a new species, S. morrisi.[11] In 2014, the species was reassigned to a new genus, Augustynolophus.[13]
Description
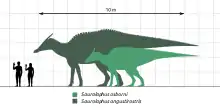
Saurolophus is known from material including nearly complete skeletons, giving researchers a clear picture of its bony anatomy. S. osborni, the rarer Albertan species, was around 8.2–8.5 m (27–28 ft) long, with its skull 1.0 m (3.3 ft) long.[14][15] It has been estimated to have weighed around 3 tonnes (3.0 long tons; 3.3 short tons).[15] S. angustirostris, the Mongolian species, was larger; it got as large as 13 m (43 ft) in length, and larger remains are reported. It has been estimated to have potentially weighed up to 11 tonnes (11 long tons; 12 short tons).[15] The largest known skull of S.angustirostris measures 1.22 m (4.0 ft) in length.[8] Aside from size, the two species are virtually identical, with differentiation hindered by lack of study.[10]
Skull
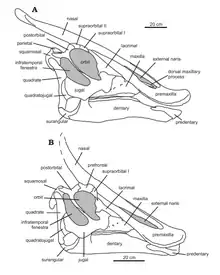
The most distinctive feature of Saurolophus is its cranial crest, which is present in young individuals, but is smaller. It is long and spike-like and projects upward and backward at about a 45° angle, starting from over the eyes. This crest is often described as solid, but appears to be solid only at the point, with internal chambers that may have had a respiratory and/or heat-regulation function.[16] The unique crest of Saurolophus is made up almost completely by the nasal bones, and in S. angustirostris it is solid. In adult specimens the crests are a rounded triangular shape in cross section. The crest protrudes past the edge of the skull backwards. Thin processes from the frontals and prefrontals extend along the underside of the crest, probably to strengthen it. At the end of the crest is a swelling of the nasal, which is often termed differently.[8]


The holotype of S. angustirostris is a skull and postcrania, so the cranium of the species is well-described. Bell et al. re-evaluated the entire species in a 2011 publication with Acta Palaeontologica Polonica. Their description found the skull to be generalized among hadrosaurines, and are much larger than any skulls of S. osborni. The most unusual feature for a hadrosaurine is the long, protruding, solid crest that extends upwards diagonally from the back of the skull roof. Unlike lambeosaurines, the crests are made up completely of the nasal bone. The premaxilla bones make up almost 50% of the entire skull length, and both sides are filled with small holes. Only in adult individuals has the front of the premaxillary contact been fused. Longer than the premaxilla, the nasal bones are the longest in the skull. They make up the entire length of the crest, and are never preserved as fused.[8]
Classification

Barnum Brown, who described the first specimens, put it in its own subfamily in "Trachodontidae" (=Hadrosauridae), the Saurolophinae. At the time, this also included Corythosaurus and Hypacrosaurus, the only well-known examples of what would become the Lambeosaurinae.[3] Brown thought that Saurolophus had an expanded tip to the ischium bone in the hip, as dinosaurs now recognized as lambeosaurines had, but this appears to have been based on a mistakenly associated lambeosaurine ischium. Additionally, he misinterpreted the crests of Saurolophus and lambeosaurines as being made of the same bones.[17]
Most publications before 2010 classified Saurolophus as a member of Hadrosaurinae, often known colloquially as the "flat-headed hadrosaurs". In 2010, the subfamily Saurolophinae was brought back into use because Hadrosaurus appears to have branched off prior to the "hadrosaurine"–lambeosaurine split. As a result, Hadrosaurinae by definition cannot include the traditional "hadrosaurines". Saurolophinae is the oldest available name for the former "hadrosaurine" clade. Saurolophus, as the name suggests, is a saurolophine, as it has a saurolophine pelvis and a (largely) solid crest.[18]
The following cladogram of hadrosaurid relationships was published in 2013 by Alberto Prieto-Márquez et al. in Acta Palaeontologica Polonica:[19]
| Saurolophinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
Feeding
As a hadrosaurid, Saurolophus would have been a bipedal/quadrupedal herbivore, eating a variety of plants. Its skull permitted a grinding motion analogous to chewing, and its teeth were continually replacing and packed into dental batteries that contained hundreds of teeth, only a relative handful of which were in use at any time. Plant material would have been cropped by its broad beak, and held in the jaws by a cheek-like organ. Its feeding range would have extended from the ground to about 4 m (13 ft) above.[7]
Crest function

The distinctive spike-like crest of Saurolophus has been interpreted in multiple ways, and could have had multiple functions. Brown compared it to the crest of a chameleon, and suggested it could provide an area for muscle attachment and a connection point for a nonbody back frill like that seen in the basilisk lizard. Peter Dodson interpreted similar features in other duckbills as having use in sexual identification.[20] Maryańska and Osmólska, noting the hollow base, suggested that the crest increased the surface area of the respiratory cavity, and helped in thermoregulation.[16] James Hopson supported a function as a visual signal, and further mentioned the possibility that the inflatable skin flaps over the nostrils could have acted as resonators and additional visual signals.[21] This idea has been picked up by authors of popular dinosaur works, such as David B. Norman, who discussed hadrosaurid display at length and included a life restoration of such an adaptation in action.[22]
Ontogeny




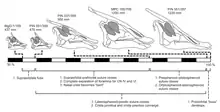
In 2015 Leonard Dewaele and colleagues described a small and partial nest containing several juveniles of S. angustirostris. The specimen (MPC-D 100/764) was recovered from the notorious Dragon's Tomb assambleage of the Nemegt Formation. The team noted that among remains, three or even four juveniles can be recognized, and two fragmentary eggshells were found in association. Juveniles within this block were identified as perinates, as they had skull lengths less than five percent of the length of the skulls of the adults, indicating they were in the earliest developmental stage at the time of their deaths. Based on these juveniles, Dewaele and team indicated that during the ontogeny of S. angustirostris the distinct crest found in adults was poorly developed in infancy, the snout grew proportionally longer, the orbit became more oval-shaped, the doming of the frontal became less prominent, and the coronoid process became higher.[23]
Social behavior
Bell and team in 2018 described the famous Dragon's Tomb assambleage of the Altan Uul II locality, Nemegt Formation, which contains a large-sized bonebed of S. angustirostris. This bonebed is largely monodominant (one dominant species), with at least three size-classes (juveniles, subadults, and adults) of S. angustirostris. Examinations made to Dragon's Tomb suggest that at least 21 Saurolophus individuals can be currently found. The team indicated that this bonebed has a minimum size of about 2000 m2, which suggest that over 100 Saurolophus carcasses may have contributed to the event. However, they discussed that even though evidence clearly reflects a catastrophic mass-mortality of a social group of S. angustirostris and provide the first evidence of gregariousness in this taxon, the exact conditions and cause surrounding the group death can not be determined. Bell and team also noted that while Dragon's Tomb provides direct evidence for social behaviour in S. angustirostris, there is yet no evidence for it in S. osborni. Nevertheless, gregariousness is apparently widespread in hadrosaurines.[24]
Paleopathology
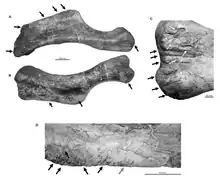
David W.E. Hone and Mahito Watabe in 2011 reported the left humerus of a nearly complete S. angustirostris skeleton (MPC-D 100/764) from the Bügiin Tsav locality of the Nemegt Formation, which was heavily damaged from bite marks attributed to the sympatric Tarbosaurus. As suggested by the lack of damage to the rest of the skeleton (such as large wounds in skeletal remains indicative of predation), this tyrannosaurid was likely scavenging an already dead S. angustirostris. It is unlikely that a large-bodied predator such as Tarbosaurus would have left sparse feeding traces on a single humerus having an entire carcass to feed on. The humerus shows three distinctive feeding methods, interpreted as punctures, drag marks, and bite−and−drag marks. Hone and Watabe noted that bite marks were mostly located at the deltopectoral crest, suggesting that this Tarbosaurus was actively selecting which biting style employ to scavenge the bone.[25]
Daily activity
Comparisons between the scleral rings of Saurolophus and modern birds and reptiles suggest that it may have been cathemeral, active throughout the day at short intervals.[26]
Paleoenvironment
Horseshoe Canyon Formation
S. osborni is known only from the upper part (unit 4) of the Horseshoe Canyon Formation. The formation is interpreted as having a significant marine influence, due to an encroaching Western Interior Seaway, the shallow sea that covered the midsection of North America through much of the Cretaceous.[27] S. osborni may have preferred to stay inland.[7] It lived alongside other dinosaur species including the ornithopods Hypacrosaurus altispinus and Parksosaurus warreni, ankylosaurid Anodontosaurus lambei, pachycephalosaurid Sphaerotholus edmontonense, ornithomimids Ornithomimus brevitertius and an unnamed species of Struthiomimus, small theropods including Atrociraptor marshalli and Albertonykus borealis, and the tyrannosaurid Albertosaurus sarcophagus.[28] The dinosaurs from this formation form part of the Edmontonian land vertebrate age.[27] A 2001 study suggested that Saurolophus osborni was part of a distinct inland fauna characterized by an association between Anchiceratops ornatus and it, while the contemporary coastal fauna was characterized by the association of Pachyrhinosaurus canadensis and Edmontosaurus regalis.[29] However, the association between S. osborni and Anchiceratops was later noted to be in error, Anchiceratops only occurs lower in the Horseshoe Canyon Formation, before the major transgression of the Western Interior Seaway represented by the Drumheller Marine Tongue.[30]
Nemegt Formation

S. angustirostris was one of the largest herbivores of the Nemegt Formation, which lacked large ceratopsians, but had sauropods and a more diverse theropod fauna. Unlike other Mongolian formations like the well-known Djadochta Formation that includes Velociraptor and Protoceratops, the Nemegt is interpreted as being well-watered region, like the Dinosaur Park Formation in Alberta.[27] When examined, the rock facies of the Nemegt formation suggest the presence of stream and river channels, mudflats, and shallow lakes. Sediments also indicate that a rich habitat existed, offering diverse food in abundant amounts that could sustain Cretaceous dinosaurs.[31] It coexisted with the rare hadrosaurid Barsboldia, flat-headed pachycephalosaurian Homalocephale and domed Prenocephale, the large ankylosaurid Saichania, rare titanosaurs sauropods Nemegtosaurus and Opisthocoelicaudia, the alvarezsaurid Mononykus, three types of troodontids including Zanabazar, several oviraptorosaurians including Rinchenia and Nemegtomaia, the ornithomimosaurs Anserimimus and Gallimimus, and the giant theropods Deinocheirus and Therizinosaurus, including the tyrannosaurid Tarbosaurus.[28]
S. angustirostris was common, and would have been an important large herbivore in the Nemegt Formation. By comparison, S. osborni was rare in the Horseshoe Canyon Formation, and faced competition from other duckbills (genus Hypacrosaurus).
See also
- Timeline of hadrosaur research
References
- Brown, Barnum (1912). "A crested dinosaur from the Edmonton Cretaceous". Bulletin of the American Museum of Natural History. 31 (14): 131–136. hdl:2246/1401.
- Brown, Barnum (1913). "The skeleton of Saurolophus, a crested duck-billed dinosaur from the Edmonton Cretaceous". Bulletin of the American Museum of Natural History. 32 (19): 387–393. hdl:2246/1415.
- Brown, Barnum (1914). "Corythosaurus casuarius, a new crested dinosaur from the Belly River Cretaceous, with provisional classification of the family Trachodontidae". Bulletin of the American Museum of Natural History. 33 (55): 559–564. hdl:2246/1734.
- Riabinin, Anatoly Nikolaenvich, N. (1930). "On the age and fauna of the dinosaur beds on the Amur River". Mémoir, Société Mineral Russia (in Russian). 59: 41–51.
- Rozhdestvensky, Anatoly K. (1952). Новый представитель утконосых динозавров из верхнемеловых отложений Монголии [A new representative of the duck-billed dinosaurs from the Upper Cretaceous deposits of Mongolia]. Doklady Akademii Nauk SSSR (in Russian). 86 (2): 405–408.
- Glut, Donald F. (1997). "Saurolophus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 788–789. ISBN 0-89950-917-7.
- Horner, John R.; Weishampel, David B.; Forster, Catherine A (2004). "Hadrosauridae". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 438–463. ISBN 0-520-24209-2.
- Bell, P. R. (2011). "Cranial Osteology and Ontogeny of Saurolophus angustirostrisfrom the Late Cretaceous of Mongolia with Comments on Saurolophus osbornifrom Canada". Acta Palaeontologica Polonica. 56 (4): 703–722. doi:10.4202/app.2010.0061.
- Bell, P.R. (2012). "Standardized Terminology and Potential Taxonomic Utility for Hadrosaurid Skin Impressions: A Case Study for Saurolophus from Canada and Mongolia". PLOS ONE. 7 (2): e31295. Bibcode:2012PLoSO...731295B. doi:10.1371/journal.pone.0031295. PMC 3272031. PMID 22319623.
- Norman, David B.; Sues, Hans-Dieter (2000). "Ornithopods from Kazakhstan, Mongolia and Siberia". In Benton, Michael J.; Shishkin, Mikhail A.; Unwin, David M.; Kurochkin, Evgenii N. (eds.). The Age of Dinosaurs in Russia and Mongolia. Cambridge: Cambridge University Press. pp. 462–479. ISBN 0-521-55476-4.
- Albert Prieto-Márquez & Jonathan R. Wagner (2013). "A new species of saurolophine hadrosaurid dinosaur from the Late Cretaceous of the Pacific coast of North America". Acta Palaeontologica Polonica. 58 (2): 255–268. doi:10.4202/app.2011.0049.
- Bell, P.R.; Evans, D.C. (2010). "Revision of the status of Saurolophus (Hadrosauridae) from California, USA". Canadian Journal of Earth Sciences. 47 (11): 1417–1426. Bibcode:2010CaJES..47.1417S. doi:10.1139/E10-062.
- Prieto-Márquez, Albert; Wagner, Jonathan R.; Bell, Phil R.; Chiappe, Luis M. (2014). "The late-surviving 'duck-billed' dinosaur Augustynolophus from the upper Maastrichtian of western North America and crest evolution in Saurolophini". Geological Magazine. 152 (2): 225–241. doi:10.1017/S0016756814000284. S2CID 131049979.
- Lull, Richard Swann; Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper 40. Geological Society of America. p. 226.
- Paul, Greg (2010). The Princeton Field Guide to Dinosaurs. New Jersey: Princeton University Press. p. 335.
- Maryańska, Teresa; Osmólska, Halszka (1981). "Cranial anatomy of Saurolophus angustirostris with comments on the Asian Hadrosauridae (Dinosauria)" (PDF). Palaeontologia Polonica. 42: 5–24.
- Sternberg, Charles M. (1954). "Classification of American duckbilled dinosaurs". Journal of Paleontology. 28 (3): 382–383.
- Prieto-Márquez, Alberto (2010). "Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods". Zoological Journal of the Linnean Society. 159 (2): 435–502. doi:10.1111/j.1096-3642.2009.00617.x.
- Prieto-Márquez, A.; Wagner, J.R. (2013). "A new species of saurolophine hadrosaurid dinosaur from the Late Cretaceous of the Pacific coast of North America". Acta Palaeontologica Polonica. 58 (2): 255–268. doi:10.4202/app.2011.0049.
- Dodson, Peter (1975). "Taxonomic implications of relative growth in lambeosaurine dinosaurs". Systematic Zoology. 24 (1): 37–54. doi:10.2307/2412696. JSTOR 2412696.
- Hopson, James A. (1975). "The evolution of cranial display structures in hadrosaurian dinosaurs". Paleobiology. 1 (1): 21–43. doi:10.1017/S0094837300002165. S2CID 88689241.
- Norman, David B. (1985). "Hadrosaurids II". The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books. pp. 122–127. ISBN 0-517-46890-5.
- Dewaele, Leonard; Tsogtbaatar, Khishigjav; Barsbold, Rinchen; Garcia, Géraldine; Stein, Koen; Escuillie, François; Godefroit, Pascal (October 14, 2015). "Perinatal specimens of Saurolophus angustirostris (Dinosauria: Hadrosauridae), from the Upper Cretaceous of Mongolia". PLOS ONE. 10 (10): e0138806. Bibcode:2015PLoSO..1038806D. doi:10.1371/journal.pone.0138806. PMC 4605499. PMID 26466354.
- Bell, P. R.; Evans, D. C.; Eberth, D. A.; Fanti, F.; Tsogtbaatar, K.; Ryan, M. J. (2018). "Sedimentological and taphonomic observations on the "Dragon's Tomb" Saurolophus (Hadrosauridae) bonebed, Nemegt Formation (Upper Cretaceous), Mongolia". Palaeogeography, Palaeoclimatology, Palaeoecology. 494: 75−90. Bibcode:2018PPP...494...75B. doi:10.1016/j.palaeo.2017.11.034.
- Hone, D. W. E.; Watabe, M. (2011). "New information on scavenging and selective feeding behaviour of tyrannosaurids" (PDF). Acta Palaeontologica Polonica. 55 (4): 627−634. doi:10.4202/app.2009.0133.
- Schmitz, L.; Motani, R. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. 332 (6030): 705–8. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. PMID 21493820. S2CID 33253407.
- Dodson, Peter (1996). The Horned Dinosaurs: A Natural History. Princeton: Princeton University Press. pp. 14–15. ISBN 0-691-05900-4.
- Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M.P.; Noto, Christopher N. (2004). "Dinosaur distribution". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 517–606. ISBN 0-520-24209-2.
- Lehman, T. M., 2001, Late Cretaceous dinosaur provinciality: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 310-328.
- Sullivan, R.M. and Lucas, S. G. (2006). "The Kirtlandian land-vertebrate "age"–faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America." Pp. 7-29 in Lucas, S. G. and Sullivan, R.M. (eds.), Late Cretaceous vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35.
- Novacek, M. (1996). Dinosaurs of the Flaming Cliffs. Bantam Doubleday Dell Publishing Group Inc. New York, New York. ISBN 978-0-385-47775-8
External links
![]() Data related to Saurolophus at Wikispecies
Data related to Saurolophus at Wikispecies
Ornithopoda | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||





.jpg.webp)