ADN ligasa
ADN ligasa es una enzima de tipo ligasa que forma enlaces covalentes entre el extremo 5’ de una cadena polinucleotídica y el extremo 3’ de otra cadena polinucleotídica. También se denomina enzima de unión de polinucleótidos.
| ligasa I, ddd, ATP-dependiente | ||||
|---|---|---|---|---|
 ADN ligasa reparando cromosoma dañado. | ||||
| Estructuras disponibles | ||||
| PDB | ||||
| Identificadores | ||||
| Símbolo | LIG1 (HGNC: 6598) | |||
| Identificadores externos |
Bases de datos de enzimas
| |||
| Número EC | 6.5.1.1 | |||
| Locus | Cr. 19 | |||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
| PubMed (Búsqueda) |
| |||
| PMC (Búsqueda) |
| |||
| ligasa III, ADN, ATP-dependiente | ||||
|---|---|---|---|---|
| Estructuras disponibles | ||||
| PDB | Buscar ortólogos: PDBe, RCSB | |||
| Identificadores | ||||
| Símbolo | LIG3 (HGNC: 6600) | |||
| Identificadores externos | ||||
| Locus | Cr. 17 q11.2-q12 | |||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
| ligasa IV, ADN, ATP-dependiente | ||||
|---|---|---|---|---|
| Estructuras disponibles | ||||
| PDB | Buscar ortólogos: PDBe, RCSB | |||
| Identificadores | ||||
| Símbolo | LIG4 (HGNC: 6601) | |||
| Identificadores externos | ||||
| Locus | Cr. 13 q33-q34 | |||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
La reparación por escisión es realizada por 3 enzimas: la correndonucleasa U.V., iniciadora del proceso, la ADN polimerasa I, que elimina y reconstruye el fragmento de ADN, y la ADN ligasa, que realiza la unión de los fragmentos nuevos y antiguos.
Para conseguir una replicación cuidadosa del ADN (ácido desoxirribonucleico), cada cadena de la doble hélice hace de molde para sintetizar la nueva cadena que tendrá una secuencia complementaria a la cadena molde. Por un lado, como las dos cadenas de la doble hélice tienen direcciones opuestas (una es 5'-3 'mientras que el otro es 3'-5') este proceso que parece sencillo, se complica ya que necesita mecanismos específicos para sintetizar las dos cadenas complementarias también en direcciones opuestas.
Por otro lado, en la doble hélice las cadenas están unidas por sus bases nitrogenadas y por tanto hay que hacer una cadena complementaria a cada cadena ya existente. Primero se tienen que separar, luego se crea la cadena complementaria y finalmente cada molde y su complementaria se deben volver a enrollar entre ellas para formar la nueva cadena de ADN. En este momento entran en acción las ADN ligasas para unir los extremos de esta nueva cadena.
¿Qué es la ADN ligasa?
La unión de los fragmentos de Okazaki (son las cadenas complementarias originadas a partir de las cadenas molde) con las cadenas complementarias necesita una enzima que la catalice. Esta enzima es la ADN ligasa.
Su función es catalizar un enlace fosfodiéster entre el grupo 3'-hidroxilo extremo de una de las cadenas de ADN y el grupo 5'-difosfato del extremo de la otra cadena de ADN. Para llevar a cabo esta reacción, la célula necesita energía porque se trata de una reacción termodinámicamente desfavorable, es decir, no es una reacción que sucede de manera espontánea.
Por un lado, en las células eucariotas y en las arqueas, la fuente de energía es el ATP (Adenosin Trifosfato). La molécula de ATP se separa en AMP (Adenosin Monofosfato) y pirofosfato para facilitar la dirección.
Por otro lado, las bacterias obtienen la energía del NAD+ (nicotinamida adenina dinucleótido) el cual se divide en AMP y NMN (mononucleótido de nicotinamida) con la misma finalidad que lo hace el ATP de eucariotas y arqueas.
La ADN ligasa no puede unir dos moléculas de ADN de cadena sencilla (ADN con una sola cadena) ni formar un ADN circular de cadena sencilla sino que su función es sellar las roturas que han tenido lugar en las moléculas de ADN de doble cadena (ADN con dos cadenas enrolladas en forma de doble hélice) en el momento de la separación de la doble hélice.
La bacteria escherichia coli es capaz de formar un enlace fosfodiéster si hay al menos algunas bases nitrogenadas de ADN de cadena sencilla con el extremo de un fragmento de ADN de doble cadena y formar pares de bases.
La ligasa codificada por el bacteriófago T4 puede unir dos fragmentos de doble hélice de extremos romos, esta capacidad se explota en la tecnología de ADN recombinante.
La manera más sencilla del ADN es aquella que tiene su extremo romo. Este tipo de moléculas terminan con un par de bases. Las moléculas con extremos romos tienen dos desventajas; la primera es que las cadenas con extremos romos tienen menos rendimiento y la segunda es que tiene más posibilidades de insertar el fragmento de ADN deseado en la dirección opuesta a la que se quiere. por otro lado, dos extremos romos siempre serán compatibles entre sí.
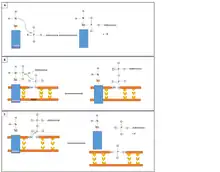
Mecanismo de actuación de las ligasa
Para formar los dos enlaces fosfodiéster covalentes entre ambos extremos de las dos cadenas, la ADN ligasa cataliza una reacción junto con el ATP que sigue 3 pasos:
- 1-Adenización de un residuo en el centro activo de la enzima liberando fosfato.
- 2- Transferencia del AMP hacia el fosfato 5 'del donante originando la formación de un enlace pirofosfato.
- 3-Formación de un enlace fosfodiéster entre el fosfato 5 'del donante y el hidroxilo 3' del aceptor.
Cuando la ligasa trabaja sobre extremos rotos requiere mayor concentración de enzimas y diferentes condiciones de reacción.
Historia
En 1967, científicos de varios laboratorios descubren la ADN ligasa. Se descubrieron originalmente en el bacteriófago T4, la bacteria escherichia coli, así como en otras bacterias.
Tipos de ADN ligasas
Existen dos ramas principales de ADN ligasas: las ADN ligasas ATP-dependientes y las NAD+-dependientes. El cofactor ATP es principal en las células de mamíferos mientras que las bacterias utilizan principalmente el NAD+ como cofactor de la enzima, aunque se encuentran bacterias con la capacidad de formar enzimas que necesitan de ATP como cofactor para funcionar. No obstante ambas enzimas siguen el mismo mecanismo reactivo.
ADN ligasas en bacterias
La ADN ligasa de la bacteria E.coli está codificada por el gen LIG. Como en muchos organismos procariotas, la ADN ligasa de esta bacteria usa la energía creada con el cofactor NAD para formar los enlaces fosfodiéster. Esta ligasa no puede unir los extremos romos de ADN excepto en ciertas condiciones moleculares de importante presencia de glicol polietileno. Tampoco puede unir de manera eficiente el ARN al ADN. Esta enzima se utiliza en laboratorios para clonar moléculas de ADN de virus dsDNa.
ADN ligasas en mamíferos
En los mamíferos, se pueden determinar hasta 4 clases de ADN ligasas cuyo substrato principal es ATP. Tres genes identificados codifican estas ADN ligasas: LIG1, LIG3 y LIG4. La ADN ligasa II al ser un derivado de la ADN ligasa III (por mecanismo proteolítico) reduce a 3 genes el número de genes implicados en la biogénesis de estas enzimas. El rol que desempeñan cada una de estas enzimas viene determinado por las proteínas con las que interacciona.
ADN ligasa I
Esta ADN ligasa es principal en el mecanismo de replicación del ADN. En el dominio del grupo amino terminal no catalítico se localiza el centro activo de esta enzima. Además la ADN ligasa I participa en la escisión del ADN así permitiendo su reparación y mantenimiento junto con otras proteínas como la beta ADN polimerasa, con la cual la ADN ligasa I interacciona de manera directa, formando así un complejo multiproteico desempeñando dicha función.
ADN ligasa II
Esta enzima es un fragmento generado por proteólisis de la ADN ligasa III.
ADN ligasa III
Dos formas de la ADN ligasa III son generadas por el gen LIG3: alfa-ADN ligasa III y beta-ADN ligasa III. Estas enzimas se diferencian por su habilidad de unirse a la proteína reparadora del ADN, XRCC1. La proteína XRCC1 solo se une a la alfa-ADN ligasa y se hace a través del dominio BRCT en el fragmento C-terminal de los polipéptidos de la ligasa. Esta proteína permite la reparación de daños en la cadena simple de ADN y la reparación por escisión de base (BER). Por otro lado la beta-ADN ligasa III parece ayudar en la finalización de la recombinación meiótica o en la reparación del ADN postmeiosis.
ADN ligasa IV
La ADN ligasa IV se une a una proteína reparadora de ADN, XRCC4. Esta unión se hace a partir de la región C-terminal de la ADN ligasa IV, que contiene dos dominios BRCT. Esta interacción, estimuladora para la actividad de unión del ADN implica que esta enzima funciona en el sistema de recombinación genética V(D)J y en la unión terminal no-homóloga de las rupturas de ADN bicatenario
Aplicaciones en la investigación de la biología molecular
Actualmente las ADN ligasas son una herramienta indispensable en la investigación, en el sector de la biología molecular, que se usa para generar secuencias de ADN recombinante.
Principalmente son usadas como enzimas de restricción para insertar fragmentos de ADN casi siempre en genes de moléculas de ADN plasmídico aunque también se puede insertar en otros.
Para realizar este tipo de experimentos se debe controlar sobre todo la temperatura. Como la mayoría de experimentos usan las ADN ligasa de T4, se realizan a una temperatura de 25 °C debido a que es la temperatura en la que la actividad enzimática de estas proteínas es mayor. Esta temperatura debe equilibrarse con la temperatura óptima de fusión para que ambas cadenas se unan. La temperatura de ligadura más eficaz para los extremos romos es de 14-20 °C.
Participación de la ADN ligasa en procesos de clonación
Para la clonación de las cadenas de ADN se utilizan vectores plasmídicos, pequeñas moléculas circulares de doble cadena, derivados de grandes plásmidos (moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientemente del ADN cromosómico) que aparecen de forma natural en casi todas las bacterias y ciertos levaduras. Como norma general suponen un pequeño fragmento del ADN total de la célula del huésped pero se pueden separar debido a su pequeño tamaño con respecto a las moléculas de ADN cromosómico de tamaño superior y que sedimentan por centrifugación.
Para poder utilizar los plásmidos circulares como vectores de clonación, el primer paso es purificarlos. Una vez se han purificados estos vectores de clonación se cortan con una nucleasa de restricción (estas nucleasas son enzimas que cortan el ADN de doble cadena cuando éstas reconocen un patrón de secuencia específico) para generar moléculas lineales. Por su parte el ADN genómico de la célula que se va a utilizar en la construcción de la genoteca es cortado con la misma nucleasa de restricción; los fragmentos de restricción resultantes (entre los que se encuentran los que contienen el ADN que va a ser clonado) se añaden a los plásmidos cortados y se cierran, formando moléculas circulares de ADN recombinante. Estas moléculas recombinantes, que contienen injertos de ADN ajeno, se unen covalentemente mediante la enzima ADN ligasa.
El siguiente paso en la preparación de la genoteca consiste en introducir las moléculas circulares de ADN recombinante en bacterias temporalmente permeables al ADN; se dice que estas células han sido transfectadas con el plásmido. A medida que estas células van creciendo y dividiéndose - duplican su número cada 30 minutos- los plásmidos recombinantes también se van replicando y produciendo un número enorme de copias de ADN circulantes que contienen ADN ajeno.
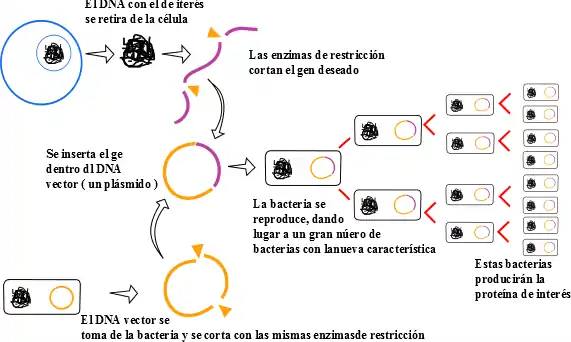
Disfunciones de la ADN ligasa
El complejo IV ligasa de ADN, que consiste en el ADN de la subunidad catalítica de ligasa IV y XRCC4 su cofactor, lleva a cabo la etapa de ligación de la reparación. XLF, también conocido como Cernunnos, es homóloga a la levadura Nej1 y también se requiere para NHEJ(“No Homologue Extreme junction" o “Extremo no homólogas de unión” es un camino que repara roturas de doble cadena en el ADN. NHEJ se conoce como "no homóloga" porque los extremos rotura se ligan directamente sin la necesidad de una plantilla homóloga, en contraste con la recombinación homóloga, lo que requiere una secuencia homóloga a la guía de reparación). Mientras que el papel preciso de XLF es desconocido, que interactúa con el complejo IV XRCC4/ADN ligasa y es probable que participa en la etapa de ligación. La evidencia reciente sugiere que XLF re-adenilatos ADN ligasa IV después de la ligadura, la recarga de la ligasa y permitiendo que catalizan una segunda ligadura.
La ADN ligasa IV y XLF se requieren para todos los eventos NHEJ.
Muchos genes NHEJ se han eliminado en los ratones. Supresión de la XRCC4 o LIG4 causas letalidad embrionaria en ratones, lo que indica que NHEJ es esencial para la viabilidad en los mamíferos. Todos los ratones NHEJ mutantes muestran un fenotipo SCID, la sensibilidad a la radiación ionizante, y la apoptosis neuronal.
La disfunción parcial de la ADN ligasa está relacionada también con Síndrome con Inestabilidad cromosómica.
Referencias
- Lehman, I. R. 1974. DNA ligase: Structure, mechanism, and function. Science 186:790-97.PubMed
- FAbO, 11-11-2007. Medicina molecular, enzima de restricción. Archivado el 20 de agosto de 2014 en Wayback Machine.
- E-Centro, 2012-2014. , Centro de Artigo, ADN-Ligasa, Art. n.º-136370.
- E-Centro, 2012-2014. , Centro de Artigo, Disfunción en NHEJ, Art. n.º-31583.
- Inmunología. Una ciencia activa 2ª edición. Juana Ángel Uribe et.al. Marzo de 2009.
- Bacterial DNA ligases. Adam Wilkinson, Jonathan Day and Richard Bowater. Molecular Microbiology (2001) 40(6), 1241–1248. Molecular Biology Sector, School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, UK.
- Viroid RNA redirects host DNA ligase 1 to act as an RNA ligase. Nohales M.Á, Flores R, Daròs J.A. 2012 Aug 21;109(34):13805-10. PMID 22869737 [PubMed - indexed for MEDLINE]
- Mammalian DNA ligases.Tomkinson AE1, Levin DS. Bioessays. 1997 Oct;19(10):893-901.PMID 9363683 [PubMed - indexed for MEDLINE]
 Datos: Q408483
Datos: Q408483 Multimedia: DNA ligase / Q408483
Multimedia: DNA ligase / Q408483