Amargasaurus cazaui
Amargasaurus cazaui ("reptil de Amarga del Dr. Luis B. Cazau") es la única especie conocida del género extinto Amargasaurus de dinosaurio saurópodo dicreosáurido, que vivió a principios del período geológico Cretácico, hace aproximadamente 130 millones de años, en el Barremiense, en lo que hoy es Sudamérica. Amargasaurus se encuentra cercanamente emparentado con el más antiguo dinosaurio africano Dicraeosaurus. Al igual que este presenta grandes espinas neurales bífidas, aunque en Amargasaurus son de mayor tamaño. El esqueleto está casi completo, incluyendo un cráneo fragmentario, haciendo de Amargasaurus uno de los saurópodos más conocidos desde el Cretácico inferior. Era pequeño para un saurópodo, alcanzando 9 a 10 metros de longitud. Su principal característica eran dos filas paralelas de altas espinas en su cuello y la espalda, más altas que en cualquier otro saurópodo conocido. No está claro si estas espinas tenían velas de la piel o sobresalían del cuerpo como estructuras solitarias con una vaina de queratina. Podrían haber sido utilizado para la visualización, combate o defensa.
| Amargasaurus cazaui | ||
|---|---|---|
| Rango temporal: 130 Ma - 122,46 Ma [1] Cretácico inferior | ||
 Réplica de un esqueleto de Amargasaurus en el vestíbulo del Museo de Melbourne. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Sauropoda | |
| Superfamilia: | Diplodocoidea | |
| Familia: | Dicraeosauridae | |
| Género: |
Amargasaurus Salgado y Bonaparte, 1991 | |
| Especie: |
A. cazaui Salgado y Bonaparte, 1991 | |
Amargasaurus fue descubierto en rocas sedimentarias de la formación La Amarga, que data entre el Barremiense y el Aptiense temprano de la época del Cretácico Inferior. Compartió su entorno con al menos otros tres géneros de saurópodos, que podría haber explotado diferentes fuentes de alimentos con el fin de reducir la competencia. Amargasaurus probablemente alimentado a media altura, como se muestra por la orientación de su oído interno y la articulación de su vértebras del cuello, lo que sugiere una posición habitual del hocico entre unos 80 centímetros y de 2,7 metros por encima del suelo. Dentro de Sauropoda, Amargasaurus está estrechamente relacionado con los géneros del Jurásico Dicraeosaurus, Brachytrachelopan y Suuwassea. Juntos, estos géneros forman la familia Dicraeosauridae, que difieren de otros saurópodos en mostrar cuellos muy cortos y pequeños tamaños de cuerpo.
Descripción

Amargasaurus era pequeño para ser saurópodo, midiendo entre 9 y 10 metros de largo[2][3][4] y una altura de 2,6 metros.[2] Habría sido un herbívoro cuadrúpedo con un cráneo largo y bajo en el extremo de un cuello relativamente corto, como en su cercano pariente Dicraeosaurus.[5] Poseía unas altas espinas, más alta que en otros saurópodos, que se bifurcaban y nacían desde las vértebras. Estas espinas se extendían desde la nuca, por el cuello hasta la mitad de la espalda. Es posible que sostuvieran una vela y suele ser representado con esta aunque esto es rechazado por Gregory S. Paul en 2000.[6] Otra posibilidad es que estuvieran recubierta de grasa o tejido muscular.
El cuerpo de Amargasaurus siguió el plan típico de un saurópodo, con una larga cola y cuello, la cabeza pequeña, y un tronco en forma de barril soportado por cuatro patas en forma de columna. El cuello de Amargasaurus era más corto que en la mayoría de otros saurópodos, un rasgo común dentro de la dicraeosauridae.[7] Midiendo de 2,4 metros de longitud,[7] el cuello corresponde a 36% de la longitud de la columna dorsal vertebral.[8] Esto es comparable a Dicraeosaurus (23%), pero mayor que el 75% en el Brachytrachelopan forma extremadamente cuello corto.[8] El cuello consistió en 13 vértebras cervicales, que eran opistocélicas, convexa en la parte delantera y el hueco en la parte posterior, articuladas en forma de bola y cavidad con vértebras vecinas.[3] El tronco poseía 9 dorsales y, probablemente, 5 vértebras sacras fusionadas.[3] Los dorsales más destacados fueron opistocélicas, mientras que los restantes fueron dorsales anfyplanas, plana en ambos extremos.[3] Las apófisis transversas robustas, proyecciones laterales que conectan con las costillas, indican una caja torácica muy desarrollado.[9] Las vértebras dorsales de Amargasaurus y otros dicraeosáuridos carecen pleurocelos, las excavaciones laterales profundos que eran característicos de otros saurópodos.[9]
La característica más llamativa del esqueleto son las extremadamente altas espinas neurales , que sobresale hacia arriba en el cuello y las vértebras dorsal anteriores. Las espinas neurales están bifurcadas a lo largo de toda su longitud, formando una fila doble. Su sección transversal era circular y la estrecha hacia sus puntas. Las espinas más altos se podían encontrar en la parte media del cuello, donde alcanzaron los 60 centímetros en la octava vértebra.[3] En el cuello, se inclinaron hacia atrás, se proyectan por encima de la vértebra adyacente.[4] En gran medida las espinas alargadas siguen a lo largo de las dos últimas vértebras dorsales, la cadera y la comienzo de la cola. Sin embargo, en estas regiones las espinas no se bifurcan pero se dilataron en un extremo superior en forma de paleta.[10] La región pélvica era relativamente amplia, a juzgar por las largas, apófisis transversas que sobresalen lateralmente de las vértebras sacras.[3] Las extremidades anteriores son algo más cortas que las patas traseras, como en saurópodos relacionados. La mayor parte de los huesos de la mano y del pie no se conservan, pero probablemente poseía cinco dígitos cada uno, como en todos los saurópodos.[9]
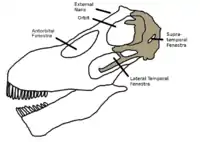
Solo se conserva la parte trasera del cráneo. Es probable que tuviese un hocico ancho como caballo equipado con dientes similares a un lápiz, como se ve en los saurópodos relacionados para los cuales se conocen cráneos más completos.[11] Como en otros dicraeosáuridos, las narinas, apertura externa de la fosa nasal, se encuentra en la mitad posterior del cráneo, en diagonal por encima de la órbita del ojo, que es proporcionalmente grande.[9][5] Al igual que en la mayoría de otros dinosaurios, el cráneo contó con tres aberturas adicionales, fenestras. La fenestra infratemporal, que se encuentra por debajo de la cuenca del ojo, era larga y estrecha.[5] Detrás de la cuenca del ojo se encuentra la fenestra supratemporal, que en los dicraeosáuridos era pequeña y se puede ver cuando el cráneo se ve desde el lado. Esto contrasta con otros reptiles diápsidos, donde estas aberturas se dirige hacia arriba, siendo así visible solo en la vista superior.[3] En frente de la cuenca del ojo, se encontraría la fenestra antorbital, aunque esta región no se conserva. Una característica inusual eran las pequeñas aberturas se ven en la parte posterior del cráneo, los llamados orificios parietales o fontanelas. En otros tetrápodos, estas aberturas se ven por lo general solo en los juveniles y cerrarían medida que el individuo crece.[12][13]
Descubrimiento e investigación
Amargasaurus es conocido por un relativamente completo único esqueleto. Este esqueleto incluye la base del cráneo, todas las vértebras del cuello, espalda y caderas, así como una porción de la cola. El hombro derecho también se conoce, al igual que el miembro superior e inferior izquierdo, y el ilion izquierdo, un hueso de la pelvis.[4] El nombre Amargasaurus fue colocado en 1991 por los paleontólogos argentinos Leonardo Salgado y Fernanda Ojeda Cruz,[9] debido de que sus restos fueron encontrados en la orilla del arroyo La Amarga en Neuquén provincia de la Patagonia Argentina. La Amarga es también el nombre de una localidad cercana y de la formación geológica donde fue encontrado. El nombre específico, A. cazaui se le dio en honor del hombre que descubrió el sitio, el Dr. Luis B. Cazau, geólogo de YPF. En 1983, Cazau informó al equipo de Bonaparte acerca del significado paleontológico de la Formación La Amarga, llevando al descubrimiento del esqueleto.[9]

El único esqueleto conocido, MACN-N 15, fue descubierto en febrero de 1984 por Guillermo Rougier durante una expedición dirigida por el famoso paleontólogo argentino José Fernando Bonaparte. Esta fue la octava expedición del proyecto "Vertebrados Terrestres del Jurásico y Cretácico de América del Sur", que fue apoyada por la National Geographic Society y se inició en 1975 con el objetivo de mejorar la, en ese momento, escaso conocimiento de tetrápodos del Jurásico y Cretácico de América del Sur.[9][14] La misma excursión descubrió el esqueleto casi completo del terópodo con cuernos Carnotaurus.[15] El sitio del descubrimiento se encuentra en el arroyo La Amarga en el Departamento Picún Leufú de la provincia de Neuquén, a unos 70 kilómetros al sur de Zapala.[9][16] el esqueleto se desenterró de rocas sedimentarias de la formación La Amarga, que data del Barremiense al Aptiense temprano, etapas del Cretácico inferior, hace más o menos alrededor de 130 a 120 millones de años.[3]

El esqueleto está razonablemente completo e incluye un cráneo parcial. Los huesos del cráneo de los saurópodo se encuentran raramente,[17] y el cráneo de Amargasaurus es solamente el segundo cráneo conocido de un Dicraeosauridae. Se encontraron las partes principales del esqueleto en su posición anatómica original, La columna vertebral del cuello y la espalda, que constaba de 22 vértebras articuladas, se encontró conectada tanto al cráneo como al sacro. Del cráneo solo se conservan la región temporal y la base del cráneo. El sacro, a pesar de haber sido parcialmente erosionado antes de haber sido encontrado, está bastante completo. Falta la mayor parte de la cola, conservándose tres vértebras anteriores, tres medias y una posterior, junto con fragmentos de varias otras. La cintura escapular se conoce por la escápula y el coracoide, que se encuentra en el extremo inferior de la escápula, mientras que la pelvis se conoce simplemente a partir del ilion. Los miembros son igualmente fragmentarios, con las manos y la mayoría de los pies desaparecidos. El esqueleto se almacena actualmente en la colección del Museo Argentino de Ciencias Naturales Bernardino Rivadavia de Buenos Aires, donde se expone una réplica.[9]
La primera mención no oficial de Amargasaurus como nuevo género de dinosaurio fue publicada por J. F. Bonaparte en el libro italiano de 1984 Sulle Orme dei Dinosauria. Aquí, la especie se designó como "Amargasaurus groeberi", que se cambió en Amargasaurus cazaui en la descripción oficial publicada varios años más tarde.[18] La descripción oficial, ha sido publicada en 1991 por Leonardo Salgado y J. F. Bonaparte en la revista científica argentina Ameghiniana.[9] Un año más tarde, Salgado y Jorge O. Calvo publicó un segundo artículo se centra en la descripción del cráneo.[5]
Clasificación
Amargasaurus y Dicraeosaurus junto con el saurópodo de cuello corto Brachytrachelopan se colocan en la familia Dicraeosauridae. Los dicreosáuridos junto con los diplodócidos son incluidos dentro de Flagellicaudata.[8][19] La discusión comparativa de esta nueva especie sugiere que es más derivada que el Dicraeosaurus hansemanni de las últimas camas jurásicas de Tendaguru, en la bifurcación más alargada y más pronunciada de las vértebras presacras, y en los procesos más largos del basiterigoideo. La comparación de los géneros Dicraeosaurus - Amargasaurus con otros clados de saurópodos, particularmente con Diplodocidae, sugiere que correspondan a una familia propia de saurópodos más derivados con diferencias significativas a Diplodocidae.[9]
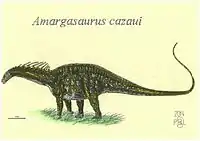
Amargasaurus está clasificado como un miembro de Dicraeosauridae, una familia clasificada clado dentro de la Diplodocoidea. En la actualidad, este clado se compone de cinco especies pertenecientes a cuatro géneros. Estos son, además Amargasaurus cazaui, las especies de Dicraeosaurus D. hansemanni y D. sattleri del Jurásico Superior en las formación Tendaguru de Tanzania y los recientemente descritos Brachytrachelopan mesai del Jurásico Tardío de Argentina. Whitlock en 20111 argumentó que Suuwassea emilieae de la formación Morrison de los Estados Unidos tiene que ser colocado dentro de Dicraeosauridae, cosa que fue apoyada por estudios posteriores.[20][21] Amargasaurus es el único dicreosáurido nombrado del cretáceo. Sin embargo, un espécimen sin nombre de Brasil indica que este grupo se mantuvo hasta el final del Cretácico Temprano.[3] La mayoría de los análisis encuentran a Dicraeosaurus y Brachytrachelopan más estrechamente relacionados entre sí que a Amargasaurus.[8][22][23] Suuwassea se recuperó como el miembro más basal de la familia.[20] En un análisis reciente de Tschopp et al. llegó al resultado preliminar de que dos géneros poco conocidos de la formación Morrison, Dyslocosaurus polyonychius y Dystrophaeus viaemalae, podrían ser miembros adicionales de Dicraeosauridae.[21] En 2016 Pablo Ariel Gallina reclasificó a Amargatitanis como un dicreosáurido.[24]
Junto con Diplodocidae y Rebbachisauridae , Dicraeosauridae está anidado dentro del Diplodocoidea. Todos los miembros de la Diplodocoidea se caracterizan por su forma de caja del hocico y los dientes estrechos restringidas a la parte más anterior de las mandíbulas. Tanto Dicraeosauridae y Diplodocidae se caracterizan por espinas neurales bifurcadas de la vértebra cervical y dorsal. En Dicraeosauridae, las espinas neurales bifurcadas estaban fuertemente alargadas, una tendencia de llegar a su extremo en Amargasaurus.[11]

El siguiente cladograma muestra el análisis filogenético de Tschopp et al. del 2015.[21]
| Dicraeosauridae |
| ||||||||||||||||||||||||||||||
Paleobiología
Espinas vertebrales

Tanto la presentación en vida de las espinas vertebrales extremadamente alargadas y bifurcadas como su función siguen siendo discutida.[3] Salgado y Bonaparte en 1991 sugirieron que las espinas fueron usadas como armas de defensa contra los depredadores, argumentando que se inclinaban hacia sus puntas.[9][13] Alternativamente, pueden haber sido utilizadas para exhibirse, ya sea para la intimidación de rivales coespecificos o para el cortejo.[9] En un dibujo de reconstrucción esquelética publicado posteriormente, Salgado en 1999 describió que las espinas neurales que soportan una vaina queratínica, lo que podría haber añadido significativamente a la longitud total de las espinas.[9][25][26] La presencia de algún tipo de recubrimiento, ya sea de queratina o piel, se muestra por un patrón estriado en la superficie de las espinas similares a los núcleos de hueso de las astas de los bovinos de hoy en día .[26]
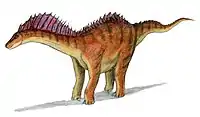
Como señaló Jack Bailey en 1997, las espinas se asemejaban a las de las velas de los pelicosaurios como Dimetrodon. Según Bailey, Amargasaurus podría también haber poseído una vela similar, que pudo haber sido utilizada para la exhibición. Además de los pelicosaurios, las espinas neurales de Amargasaurus se bifurcaban, formando una fila doble a lo largo del cuello y la espalda. Como el espacio entre ambas filas era de solo 3 a 7 centímetros, la existencia de dos velas paralelas parece poco probable. En cambio, Bailey sugiere que las espinas representan un andamio que estaba completamente envuelto por una sola capa de piel.[10] El escritor de ciencia estadounidense Gregory Paul en el 2000 argumentó en contra de la hipótesis de la vela, observando que las velas del cuello habrían reducido la flexión del cuello y que las espinas eran de sección circular en lugar de aplanadas, como ocurre en los animales que llevan vela. Las espinas, si eran alargadas por una vaina queratínica, podrían haber sido utilizadas como arma tanto contra los depredadores como contra los congéneres, ya que el animal podría haber apuntado sus espinas más anteriores hacia adelante inclinando su cuello. También presentó la hipótesis de que las espinas podrían haber sido aplaudidas entre sí para generar sonido.[25] En 2022 se comprobó la hipótesis qué Amargasaurus tenía una vela, aun no se sabe cual habría sido su función correcta.
Daniela Schwarz et al. en 2007 concluyeron que las espinas neurales bifurcadas de los diplodocidos y dicreosauridos encerraban un saco aéreo, que habría sido conectado a los pulmones como parte del sistema respiratorio. En Dicraeosaurus, este saco aéreo, el denominado divertículo supravertebral, habría descansado sobre el arco neural y llenado todo el espacio entre las espinas. En Amargasaurus, sin embargo, los dos tercios superiores de las espinas se habrían cubierto por una vaina de queratina, restringiendo el saco de aire al espacio al tercio inferior de las espinas.[26]
Las espinas neurales desde la penúltima vértebra dorsal hasta las vértebras anteriores de la cola todavía eran fuertemente alargadas, pero de estructura diferente, formando una sola fila de proyecciones en forma de paletas. Según Bailey, estas proyecciones se parecían a las de los ungulados jorobados modernos como el bisonte, lo que indica la presencia de una joroba carnosa por encima de las caderas. Bailey sugirió jorobas similares para otros dinosaurios con espinas neurales fuertemente alargadas, como Spinosaurus y Ouranosaurus.[10]
Posición del cráneo y sentidos
Paulina Carabajal et al. realizaron una exploración CT del cráneo en 2014, generando modelos tridimensionales tanto del molde de la cavidad cerebral como del oído interno. Usando estos modelos, se demostró que la cavidad craneana abarcaba de 94 a 98 mililitros en volumen. El oído interno tenía 30 milímetros de alto y 22 milímetros de ancho. La lagena, la parte que contiene las células ciliadas para la audición, era bastante corta, lo que indica que el sentido de la audición habría sido más pobre en Amargasaurus que en otros saurópodos para los que se han estudiado los oídos internos.[28]

Las primeras reconstrucciones esqueléticas muestran el cráneo en una postura casi horizontal. Salgado en 1999 argumentó que tal postura habría sido anatómicamente imposible debido a las espinas neurales alargadas de las vértebras del cuello. En su lugar, contempló la cabeza en una orientación casi vertical.[12] La orientación habitual de la cabeza suele reflejarse por la orientación de los canales semicirculares del oído interno, que albergaba al sistema vestibular responsable del sentido del equilibrio. Utilizando su modelo tridimensional del oído interno, Carabajal et al. sugirieron que el hocico se dirigía hacia abajo en un ángulo de aproximado de 65 ° con respecto a la horizontal.[28] Recientemente se ha propuesto un valor similar para Diplodocus.[23] La postura neutral del cuello puede ser aproximada basada en cómo las vértebras cervicales se unen entre sí. Según Carabajal et al., el cuello se inclinaba suavemente hacia abajo, de modo que el hocico hubiera descansado unos 80 cm por encima del suelo en postura neutra. En realidad, la postura del cuello habría variado según las respectivas actividades de los animales. El alzar del cuello, para alcanzar una posición de alerta, habría sido constreñida por las espinas neurales alargadas, no permitiendo alturas mayores de 270 cm.[28]
Locomoción

Amargasaurus era cuadrúpedo y probablemente incapaz de levantarse en sus patas traseras.[9] Salgado y Bonaparte en 1991 sugirieron que Amargasaurus era un andador lento, ya que los antebrazos y las piernas eran proporcionalmente cortos, una característica común a los animales de movimiento lento.[9] Esto fue contradicho por Gerardo Mazzetta y Richard Fariña en 1999, que argumentaron que Amargasaurus era capaz de una locomoción rápida. Durante la locomoción, los huesos de las piernas están fuertemente afectados por momentos de flexión, que representan por lo tanto un factor limitante para la velocidad máxima de un animal. Los huesos de la pierna de Amargasaurus eran aún más resistentes que los del rinoceronte blanco actual, lo que se adapta al galope.[29]
Paleoecología
Amargasaurus proviene de las rocas sedimentarias de la formación La Amarga, que forma parte de la Cuenca Neuquina y data del Barremiense y Aptiense tardío del Cretácico Inferior. La mayoría de los fósiles de vertebrados, incluyendo Amargasaurus, se han encontrado en la parte más baja, más antigua, de la formación, el miembro Puesto Antigual. Este miembro tiene aproximadamente 29 metros de espesor y está compuesto principalmente de areniscas depositadas por ríos trenzados.[30] El propio esqueleto de Amargasaurus se recuperó de una capa compuesta de conglomerados arenosos.[9] La fauna de saurópodos de la formación La Amarga fue diversa e incluyó el rebaquisáurido basal Zapalasaurus, el también dicreosáurido Amargatitanis,[24] y restos sin nombre de titanosauriformes basales.[31] La gran diversidad sugiere que diferentes especies de saurópodos explotaron diferentes fuentes de alimentos para reducir la competencia. Los titanosauriformes basales mostraron un cuello proporcionalmente más largo, patas delanteras más largas, y coronas dentales más anchas que los dicreosáuridos y rebaquisáuridos, sugiriendo mayores alturas de alimentación. Amargasaurus probablemente se alimentado por encima del nivel del suelo en alturas de hasta 2,7 metros, como lo demuestra la anatomía de su cuello y el oído interno. Rebaquisáuridos como Zapalasaurus presumiblemente se alimentaba a nivel del suelo, mientras que los titanosauriformes basales explotaron fuentes de alimentos en niveles más altos.[28]

Otros dinosaurios de la formación La Amarga incluyen el estegosaurio Amargastegos,[32] los dinosaurios depredadores incluyen el pequeño ceratosaurio Ligabueino, y la presencia de un tetanuro grande conocido solo por los dientes. Aparte de los dinosaurios, la formación es famosa por el mamífero cladoterico Vincelestes, el único mamífero conocido desde el Cretácico Inferior de América del Sur.[31] Cocodrilomorfos están presentes con el trematochámpsido Amargasuchus, cuyo holotipo fue encontrado en la asociación con los huesos de Amargasaurus.[16]
Referencias
- «Amargasaurus». Paleobiology Database (en inglés). Archivado desde el original el 24 de noviembre de 2015. Consultado el 30 de agosto de 2015.
- Mazzetta, G. V.; P. Christiansen; R. A. Farina (2004). «Giants and bizarres: body size of some southern South American Cretaceous dinosaurs». Historical Biology 16 (2–4): 71-83. doi:10.1080/08912960410001715132.
- Novas, Fernando E. (2009). The age of dinosaurs in South America. Bloomington: Indiana University Press. ISBN 978-0-253-35289-7.
- Upchurch, P., Barrett, P.M, & Dodson, P. 2004. Sauropoda. In: Weishampel, D.B., Dodson, P., & Osmolska, H. (Eds.). The Dinosauria (2nd Edition). Berkeley: University of California Press. Pp. 259–322.
- Salgado, L. & Calvo, J.O. 1992. Cranial osteology of Amargasaurus cazaui Salgado and Bonaparte (Sauropoda, Dicraeosauridae) from the Neocomian of Patagonia. Ameghiniana 29: 337-346.
- Paul, Gregory S. (2000) The Scientific American Book of Dinosaurs, p 94. St. Martin's Press. ISBN 0-312-26226-4.
- Senter, Phil (1 de enero de 2007). «Necks for sex: sexual selection as an explanation for sauropod dinosaur neck elongation». Journal of Zoology 271 (1): 45-53. doi:10.1111/j.1469-7998.2006.00197.x.
- Rauhut, OWM; Remes, K; Fechner, R; Cladera, G; Puerta, P (2005). «Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia». Nature 435 (7042): 670-672. Bibcode:2005Natur.435..670R. PMID 15931221. doi:10.1038/nature03623.
- Salgado L & Bonaparte JF. (1991). Un nuevo saurópodo Dicraeosauridae, Amargasaurus cazaui gen. et sp. nov., de la Provincia del Neuquén, Argentina. Ameghiniana 28: 333-346.
- Bailey, Jack Bowman (1997). «Neural spine elongation in dinosaurs: sailbacks or buffalo-backs?». Journal of Paleontology 71 (6): 1124-1146. JSTOR 1306608.
- Wilson, Jeffrey (2005). «Overview of Sauropod Phylogeny and Evolution». En Rogers, Kristina Curry; Wilson, Jeffrey, ed. The Sauropods: Evolution and Paleobiology. University of California Press. ISBN 0-520-24623-3.
- Salgado, Leonardo (1999). «The macroevolution of the Diplodocimorpha (Dinosauria; Sauropoda): A developmental model». Ameghiniana 36 (2): 203-216.
- Salgado, Leonardo; Coria, Rodolfo A. (2005). «Sauropods of Patagonia: Systematic Update and Notes on Global Sauropod Evolution». Thunder-Lizards: The Sauropodomorph Dinosaurs. pp. 430–453. ISBN 0253345421.
- Bonaparte, José F. (1981). «Jurassic and Cretaceous Terrestrial Vertebrates of South America». National Geographic Society Research Reports, 1975 Projects: 115-125.
- Bonaparte, José F.; Novas, Fernando E.; Coria, Rodolfo A. (1990). «Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia». Contributions in Science (Natural History Museum of Los Angeles County) 416. Archivado desde el original el 21 de julio de 2010. Consultado el 1 de noviembre de 2016.
- Chiappe, Luis M. (1988). «A new trematochampsid crocodile from the Early Cretaceous of north-western Patagonia, Argentina and its palaeobiogeographical and phylogenetic implications». Cretaceous Research 9 (4): 379-389. doi:10.1016/0195-6671(88)90009-2.
- Chure, Daniel; Britt, Brooks B.; Whitlock, John A.; Wilson, Jeffrey A. (2010). «First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition». Naturwissenschaften 97 (4): 379-391. Bibcode:2010NW.....97..379C. ISSN 0028-1042. PMC 2841758. PMID 20179896. doi:10.1007/s00114-010-0650-6.
- Glut, D. F. (1997). «Amargasaurus». Dinosaurs, the encyclopedia. McFarland & Company, Inc. Publishers. pp. 121–124. ISBN 978-0-375-82419-7.
- Taylor MP & Naish D. 2005. The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda). PaleoBios. 25(2): 1-7.
- Whitlock, John A. (2011). «A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda)». Zoological Journal of the Linnean Society 161 (4): 872-915. ISSN 1096-3642. doi:10.1111/j.1096-3642.2010.00665.x.
- Tschopp, Emanuel; Mateus, Octávio; Benson, Roger B.J. (2015). «A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)». PeerJ 3: e857. PMC 4393826. PMID 25870766. doi:10.7717/peerj.857.
- Taylor, MP; Naish, D (2005). «The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda)». PaleoBios 25 (2): 1-7.
- Sereno, PC; Wilson, JA; Witmer, LM; Whitlock, JA; Maga, A (2007). «Structural Extremes in a Cretaceous Dinosaur». PLoS ONE 2 (11): e1230. Bibcode:2007PLoSO...2.1230S. PMC 2077925. PMID 18030355. doi:10.1371/journal.pone.0001230.
- Pablo Ariel Gallina. Reappraisal Of The Early Cretaceous Sauropod Dinosaur Amargatitanis Macni (Apesteguía, 2007), From Northwestern Patagonia, Argentina. Cretaceous Research (advance online publication)doi:10.1016/j.cretres.2016.04.002http://www.sciencedirect.com/science/article/pii/S0195667116300507
- Paul, Gregory S. (2000). «Restoring the Life Appearance of Dinosaurs». En Paul, Gregory S., ed. The Scientific American Book of Dinosaurs. St. Martin's Press. p. 94. ISBN 0-312-26226-4.
- Schwarz, Daniela; Frey, Eberhard; Meyer, Christian A. (2007). «Pneumaticity and soft-tissue reconstructions in the neck of diplodocid and dicraeosaurid sauropods». Acta Palaeontologica Polonica 52 (1).
- «Amargasaurus 2022».
- Paulina Carabajal, Ariana; Carballido, José L.; Currie, Philip J. (2014). «Braincase, neuroanatomy, and neck posture of Amargasaurus cazaui (Sauropoda, Dicraeosauridae) and its implications for understanding head posture in sauropods». Journal of Vertebrate Paleontology 34 (4): 870-882. doi:10.1080/02724634.2014.838174.
- Mazzetta, Gerardo V.; Farina, Richard A. (1999). «Estimacion de la capacidad atlética de Amargasaurus cazaui Salgado y Bonaparte, 1991, y Carnotaurus sastrei Bonaparte, 1985 (Saurischia, Sauropoda-Theropoda)». XIV jornadas Argentinas de paleontologia de vertebrados, Ameghiniana 36 (1): 105-106.
- Leanza, Héctor A; Apesteguı́a, Sebastián; Novas, Fernando E; de la Fuente, Marcelo S (2004). «Cretaceous terrestrial beds from the Neuquén Basin (Argentina) and their tetrapod assemblages». Cretaceous Research 25 (1): 61-87. ISSN 0195-6671. doi:10.1016/j.cretres.2003.10.005.
- Apesteguía, Sebastián (2007). «The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina)». Gondwana Research 12 (4): 533-546. doi:10.1016/j.gr.2007.04.007.
- Ulansky, R.E. (2014). «Dinosaurs Classification. Basal Thyreophora & Stegosauria». Dinologia: 1-8.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una galería multimedia sobre Amargasaurus cazaui.
Wikimedia Commons alberga una galería multimedia sobre Amargasaurus cazaui. Wikispecies tiene un artículo sobre Amargasaurus cazaui.
Wikispecies tiene un artículo sobre Amargasaurus cazaui.- Amargasaurus en Proyectodino
- Amargasaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Amargasaurus dentro de Diplodocoidea en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q311282
Datos: Q311282- Multimedia: Amargasaurus / Q311282
- Especies: Amargasaurus