Bacteriófago MS2
El bacteriófago MS2 es un virus icosaédrico que pertenece a la especie Escherichia virus MS2.[1][2] Pertenece al grupo IV de la clasificación de Baltimore (virus ARN monocatenario positivo). Infecta a la bacteria Escherichia coli y a otros miembros de la familia Enterobacteriaceae.[3] Este virus pertenece a la clase Leviviricetes, que incluye a otros bacteriófagos relacionados como el Qβ o el BZ13.[4]
| Bacteriófago MS2 | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Dominio: | Riboviria | |
| Reino: | Orthornavirae | |
| Filo: | Lenarviricota | |
| Clase: | Leviviricetes | |
| Orden: | Norzivirales | |
| Familia: | Fiersviridae | |
| Género: | Emesvirus | |
| Especie: | Escherichia virus MS2 | |
| Clasificación de Baltimore | ||
| Grupo: | IV (Virus ARN monocatenario positivo) | |
Historia
En 1961, el bacteriófago MS2 fue aislado por Alvin John Clark y se observó que era muy similar al bacteriófago f2, hoy en día miembro de la misma especie.[2][5]
En 1976, el genoma de este virus fue el primero en ser completamente secuenciado.[6] Walter Fiers y su equipo fueron los responsables de este hito, que se añadió a la primera secuenciación de un gen completo en 1972, la proteína de la cápside de este mismo virus.[7] Estas secuencias se determinaron a nivel del ARN, a diferencia de la secuenciación del genoma del bacteriófago ΦX174 en 1977, que se hizo con ADN.[8] Posteriormente, se realizaron estudios estadísticos del genoma del bacteriófago MS2 en el que se buscaron patrones en la secuencia de nucleótidos. Se encontraron varias secuencias de ADN no codificante cuya función era desconocida en 1979, cuando se desarrollaron estas investigaciones.[9]
Virología
Genoma y productos genéticos
El genoma de MS2 es uno de los más pequeños conocidos, con 3569 nucleótidos de ARN monocatenario.[6] Codifica solo cuatro proteínas: la proteína de maduración (proteína A), la proteína de lisis, la proteína de la cápside y la proteína replicasa.[3] El gen que codifica la proteína de lisis (lys) se solapa con el extremo 3' del gen anterior (cp) y el extremo 5' del gen posterior (rep), y fue uno de los primeros ejemplos conocidos de genes superpuestos. La molécula de ARN positivo sirve también como ARN mensajero (ARNm), y se traduce tras la decapsidación del virus dentro de la célula huésped. Aunque las cuatro proteínas están codificadas por el mismo ARNm, no todas se expresan en los mismos niveles; la expresión de estas proteínas está regulada por una interacción compleja entre la traducción y la estructura secundaria del ARN.

| Gen | Número de nucleótidos | Producto génico | Número de aminoácidos |
|---|---|---|---|
| mat (MS2g1)[10] | 1487 | Proteína de maduración[11] | 393 |
| cp (MS2g2)[12] | 510 | Proteína de la cápside[13] | 130 |
| lys (MS2g3)[14] | 295 | Proteína de lisis[15] | 75 |
| rep (MS2g4)[16] | 2055 | ARN replicasa, cadena beta[17] | 545 |
Estructura de la cápside

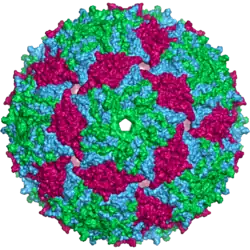
Un virión del fago MS2 tiene unos 27 nm de diámetro según los datos aportados al observarlo al microscopio electrónico.[18] La cápside está compuesta por una copia de la proteína de maduración y 180 copias de la proteína de la cápside (organizada en noventa dímeros) que forman un icosaedro con un número T de tres. Su función es proteger el ARN de su interior.[19] El virión tiene un punto isoeléctrico de 3,9.[20]
La estructura de la proteína de la cápside consta de una lámina beta de cinco cadenas con dos alfa hélices y una horquilla beta. Cuando la cápside se ensambla, las hélices y la horquilla se orientan hacia el exterior de la partícula viral, mientras que las láminas beta miran al interior.[21]

Ciclo reproductivo
El bacteriófago MS2 infecta a enterobacterias que poseen el factor de fertilidad, un plásmido que permite a las células donar ADN en la conjugación bacteriana. Los genes de este factor permiten la producción de un pilus F, que el virus utiliza como receptor. El bacteriófago se une al lateral de esta estructura con su proteína de maduración. El mecanismo por el que el ARN entra a la bacteria es desconocido.
Una vez que el ARN viral ha penetrado en la célula, comienza a funcionar como un ARN mensajero para traducir las proteínas del virus. El gen de la proteína más numerosa, la proteína de la cápside, puede ser traducido inmediatamente. El inicio del proceso de producción de la replicasa requiere cambios en la estructura secundaria del ARN, pero pueden ocurrir transitoriamente al pasar el ribosoma sobre el gen cp. Una vez que se han obtenido grandes cantidades de la proteína de la cápside, las cuales se unen a la horquilla del ARN mensajero y estabilizan su estructura, se detiene la traducción de la replicasa. La secuencia de inicio del gen mat está oculta en la estructura secundaria del ácido nucleico, pero puede exponerse durante el proceso de replicación del ARN. De esta forma solo se traducirán unas pocas copias de este gen por copia de ARN. Por último, la proteína de lisis solo puede ser producida por ribosomas que han completado la traducción del gen cp y se deslizan de vuelta a la secuencia de inicio del gen lys, algo que ocurre con una frecuencia del 5 %.[3]
La replicación de la hebra de sentido positivo de ARN requiere la síntesis de la hebra complementaria, es decir, de sentido negativo, la cual puede ser usada como moleda para la síntesis una nueva hebra positiva. Los detalles de la replicación del bacteriófago MS2 no son tan conocidos como los de fagos relacionados como el Qβ, en parte debido a que la replicasa es difícil de aislar, aunque probablemente sea similar.[3]
Se cree que la formación del virión se inicia con la unión de la proteína de maduración al ARN viral. De hecho, el complejo formado por estos dos elementos es infeccioso. El ensamblaje de las proteínas de la cápside puede ocurrir en ausencia de ARN; sin embargo, ocurre a concentraciones mucho menores cuando este está presente, porque la unión de los dímeros de proteínas de la cápside a la horquilla del ARN facilita el proceso.[3]
La lisis bacteriana y la liberación de los viriones recién formados ocurre cuando se alcanza una concentración suficiente de proteína de lisis. Esta forma poros en la membrana plasmática y provoca la pérdida del potencial de membrana y la rotura de la pared celular.[3] Se sabe que la proteína de lisis se une a la chaperona DnaJ mediante un residuo P330.[22] En todo el género Levivirus existe un dipéptido de leucilserina en la proteína de lisis, que parece ser esencial en la actividad lítica, aunque sus diferentes localizaciones sugieren que ha evolucionado de manera independiente.[23]
Aplicaciones
Desde 1998, se ha usado un método basado en el uso de horquillas de ARN de este virus y su interacción con la proteína verde fluorescente para identificar ARN en organismos vivos.[24] Otra de sus posibles aplicaciones sería usar la cápside del virus para administrar fármacos, obtener imágenes médicas de tumores y captación de luz.[25]
Se ha usado el fago MS2 en estudios sobre la transmisión de enfermedades debido a su parecido estructural con los norovirus, su capacidad similar para proliferar en ciertas condiciones y no ser capaces de producir enfermedad en humanos.[26]
Referencias
- «ICTV Taxonomy history: Escherichia virus MS2». ICTV (en inglés). Consultado el 22 de febrero de 2020.
- «Escherichia virus MS2». NCBI (en inglés). Consultado el 22 de febrero de 2020.
- van Duin, J; Tsareva, N (2006). «Single-stranded RNA phages. Chapter 15». En Calendar, R. L., ed. The Bacteriophages (2.ª edición). Oxford University Press. pp. 175–196. ISBN 978-0195148503.
- Ni, Chao-Zhou; White, Carol A.; Mitchell, Richard S.; Wickersham, John; Kodandapani, Ramadurgam; Peabody, David S.; Ely, Kathryn R. (diciembre de 1996). «Crystal structure of the coat protein from the GA bacteriophage: Model of the unassembled dimer». Protein Science (en inglés) 5 (12): 2485-2493. PMC 2143325. PMID 8976557. doi:10.1002/pro.5560051211.
- «National Academy of Sciences: Abstracts of Papers Presented at the Autumn Meeting, 29 October, La Jolla, California, 30 October-1 November 1961, Los Angeles». Science (en inglés) 134 (3488): 1425-37. November 1961. Bibcode:1961Sci...134.1425.. PMID 17795773. doi:10.1126/science.134.3488.1425.
- Fiers, W.; Contreras, R.; Duerinck, F.; Haegeman, G.; Iserentant, D.; Merregaert, J.; Min Jou, W.; Molemans, F.; Raeymaekers, A.; Van den Berghe, A.; Volckaert, G.; Ysebaert, M. (abril de 1976). «Complete nucleotide sequence of bacteriophage MS2 RNA: primary and secondary structure of the replicase gene». Nature (en inglés) 260 (5551): 500-507. Bibcode:1976Natur.260..500F. PMID 17795773. doi:10.1038/260500a0.
- Jou, W. Min; Haegeman, G.; Ysebaert, M.; Fiers, W. (mayo de 1972). «Nucleotide Sequence of the Gene Coding for the Bacteriophage MS2 Coat Protein». Nature (en inglés) 237 (5350): 82-88. Bibcode:1972Natur.237...82J. PMID 4555447. doi:10.1038/237082a0.
- Sanger, F.; Air, G. M.; Barrell, B. G.; Brown, N. L.; Coulson, A. R.; Fiddes, J. C.; Hutchison, C. A.; Slocombe, P. M. et al. (febrero de 1977). «Nucleotide sequence of bacteriophage φX174 DNA». Nature (en inglés) 265 (5596): 687-695. Bibcode:1977Natur.265..687S. PMID 870828. doi:10.1038/265687a0.
- Erickson, John W.; Altman, Gary G. (1979). «A search for patterns in the nucleotide sequence of the MS2 genome». Journal of Mathematical Biology (en inglés) 7 (3): 219-230. doi:10.1007/BF00275725. Consultado el 23 de febrero de 2020.
- «mat hypothetical protein [ Enterobacteria phage MS2 ]». NCBI (en inglés).
- «assembly protein [Enterobacteria phage MS2]». NCBI (en inglés).
- «cp hypothetical protein [ Enterobacteria phage MS2 ]». NCBI (en inglés).
- «coat protein [Enterobacteria phage MS2]». NCBI (en inglés).
- «lys hypothetical protein [ Enterobacteria phage MS2 ]». NCBI (en inglés).
- «lysis protein [Enterobacteria phage MS2]». NCBI (en inglés).
- «rep hypothetical protein [ Enterobacteria phage MS2 ]». NCBI (en inglés).
- «rna replicase beta chain [Enterobacteria phage MS2]». NCBI (en inglés).
- Strauss, James H.; Sinsheimer, Robert L. (julio de 1963). «Purification and properties of bacteriophage MS2 and of its ribonucleic acid». Journal of Molecular Biology (en inglés) 7 (1): 43-54. PMID 13978804. doi:10.1016/S0022-2836(63)80017-0.
- Valegård, Karin; Liljas, Lars; Fridborg, Kerstin; Unge, Torsten (mayo de 1990). «The three-dimensional structure of the bacterial virus MS2». Nature (en inglés) 345 (6270): 36-41. Bibcode:1990Natur.345...36V. PMID 2330049. doi:10.1038/345036a0.
- Dowd, Scot E.; Pillai, Suresh D.; Wang, Sookyun; Corapcioglu, M. Yavuz (1998). «Delineating the Specific Influence of Virus Isoelectric Point and Size on Virus Adsorption and Transport through Sandy Soils». Applied and Environmental Microbiology (en inglés) 64 (2): 405-410. PMID 9464373. doi:10.1128/aem.64.2.405-410.1998.
- Golmohammadi, Roshah; Valegård, Karin; Fridborg, Kerstin; Liljas, Lars (diciembre de 1993). «The Refined Structure of Bacteriophage MS2 at 2·8 Å Resolution». Journal of Molecular Biology 234 (3): 620-639. PMID 8254664. doi:10.1006/jmbi.1993.1616.
- Chamakura, Karthik R.; Tran, Jennifer S.; Young, Ry; Silhavy, Thomas J. (15 de junio de 2017). «MS2 Lysis of Escherichia coli Depends on Host Chaperone DnaJ». Journal of Bacteriology (en inglés) 199 (12). PMC 5446614. PMID 28396351. doi:10.1128/JB.00058-17.
- Chamakura, Karthik R.; Edwards, Garrett B.; Young, Ry (1 de julio de 2017). «Mutational analysis of the MS2 lysis protein L». Microbiology (en inglés) 163 (7): 961-969. PMC 5775895. PMID 28691656. doi:10.1099/mic.0.000485.
- Bertrand, Edouard; Chartrand, Pascal; Schaefer, Matthias; Shenoy, Shailesh M.; Singer, Robert H.; Long, Roy M. (octubre de 1998). «Localization of ASH1 mRNA Particles in Living Yeast». Molecular Cell (en inglés) 2 (4): 437-445. PMID 9809065. doi:10.1016/S1097-2765(00)80143-4.
- Glasgow, Jeff; Tullman-Ercek, Danielle (10 de mayo de 2014). «Production and applications of engineered viral capsids». Applied Microbiology and Biotechnology (en inglés) 98 (13): 5847-5858. PMID 24816622. doi:10.1007/s00253-014-5787-3.
- Fox, Maggie (8 de septiembre de 2014). «Viruses spread 'like crazy' in an office, study finds». The Today Show.
Enlaces externos
| Control de autoridades |
|
|---|
 Datos: Q106960351
Datos: Q106960351