Célula T naive
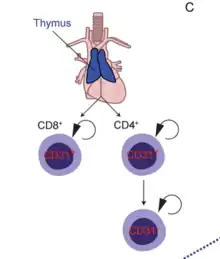
Un célula T naive, (Th0), T naive, linfocito T inexperto o linfocito T virgen es una célula T que se ha diferenciado en la médula ósea y luego ha experimentado con éxito los procesos positivos y negativos de selección central en el timo. Entre estas se encuentran las formas inexpertas de células T auxiliares (CD4+) y de células T citotóxicas (CD8+). Una célula T naive se considera madura y, a diferencia de las células T activadas o de memoria, no ha encontrado su antígeno afín en la periferia.[1]
Las células T naïve, tienen la capacidad de responder a nuevos antígenos. El número de células T naive y su diversidad (clones), representan el potencial del sistema de inmunidad adquirida para detectar y responder a patógenos nuevos y a proteínas mutantes de células malignas.[2]
Características
En los seres humanos, el conjunto de las células T naive se establece en la primera década de la vida, con la salida masiva del timo de miles de millones de células T naive recién producidas.[2]
Fenotipo
Las células T naive, se caracterizan comúnmente por la expresión superficial de L-selectina (CD62L) y el receptor de quimiocina CC tipo 7 (CCR7); la ausencia de los marcadores de activación CD25, CD44 o CD69; y la ausencia de memoria CD45RO isoforma.[3] [4] También expresan receptores funcionales de IL-7, que consisten en subunidades IL-7 receptor-a, CD127, y la cadena γ común, CD132. En el estado inmaduro, se cree que las células T requieren las citocinas de cadena gamma común IL-7 e IL-15 para los mecanismos de supervivencia homeostáticos.
Si bien las células T naive se consideran regularmente como una población de células sincronizadas en el desarrollo y bastante homogéneas y quiescentes, que solo difieren en la especificidad del receptor de células T, hay una creciente evidencia de que las células T inmaduras son realmente heterogéneas en fenotipo, función, dinámica y diferenciación estado, lo que resulta en un espectro completo de células naive con diferentes propiedades. Por ejemplo, algunas células T no inmaduras expresan marcadores de superficie similares a las células T naive (Tscm, células T de memoria de células madre;[5] Tmp, células T de memoria con un fenotipo virgen[6]), algunas células T naive de antígeno tienen perdieron su fenotipo inmaduro,[7] y algunas células T se incorporan dentro del fenotipo ingenuo de células T, pero son un subconjunto diferente de células T (Treg, células T reguladoras; RTE, emigrante tímico reciente). Es importante apreciar estas diferencias al evaluar las células T naive.
La mayoría de las células T vírgenes humanas se producen muy temprano en la vida cuando el timo del bebé es grande y funcional. La disminución en la producción de células T naive debido a la involución del timo con la edad se compensa con la llamada "proliferación periférica" o "proliferación homeostática" de células T naive que han emigrado del timo más temprano en la vida. La proliferación homeostática provoca un cambio en la expresión de genes de células T vírgenes y es decir, se manifiesta por la adquisición de la expresión de la proteína de superficie CD25.
Función
Las células T naive, pueden responder a nuevos patógenos que el sistema inmunitario aún no ha encontrado. El reconocimiento por parte de un clon virgen de células T de su antígeno afín, da como resultado el inicio de una respuesta inmune. A su vez, esto da como resultado que la célula T adquiera un fenotipo activado, que es visible por la regulación positiva de los marcadores de superficie CD25+, CD44+, CD62Lbajo, CD69+ y puede diferenciarse aún más en células T de memoria.
Tener un número adecuado de células T naive es esencial, para que el sistema inmunitario responda continuamente a patógenos desconocidos.
Mecanismo de activación
Cuando un antígeno reconocido se une al receptor de antígeno de linfocitos T (TCR) ubicado en la membrana celular de las células Th0 o naive, estas células se activan a través de la siguiente cascada de transducción de señales [8]
- "vía clásica"
- la tirosina quinasa Lck, que está asociada con los co-receptores CD4 y CD8:[9] se dedica a fosforilar el complejo coreceptor CD3 y las cadenas ζ del TCR y reclutar y activar la proteína Zap70 asociada a la cadena ζ
- Zap70 activado a su vez fosforila el adaptador de membrana Lat, que posteriormente recluta varias proteínas que contienen el dominio de homología Src, incluida la fosfolipasa C-γ1 (PLC-γ1)
- la activación de PLC-γ1 produce la hidrólisis del fosfatidilinositol 4,5-bisfosfato a inositol 3,4,5-trifosfato y diacilglicerol
- el inositol 3,4,5-trifosfato desencadena la liberación de Ca2+ de las reservas intracelulares y el diacilglicerol activa la proteína quinasa C y RasGRP
- RasGRP a su vez activa la cascada de proteína quinasa activada por mitógeno
- vía "no clásica"
implica que Zap70 activado fosforila directamente la p38 MAPK que a su vez induce la expresión del receptor de vitamina D (VDR). Además, la expresión de PLC-γ1 depende de VDR activado por calcitriol.[8] Las células T inmaduras tienen una expresión muy baja de VDR y PLC-γ1. Sin embargo, la señalización de TCR activada a través de p38 aumenta la expresión de VDR y el VDR activado con calcitriol a su vez aumenta la expresión de PLC-γ1. Por lo tanto, la activación de las células T inmaduras depende crucialmente de niveles adecuados de calcitriol.
La activación de los linfocitos T primero requiere activación a través de la ruta no clásica, para aumentar la expresión de VDR y PLC-γ1 antes de que pueda continuar la activación a través de la ruta clásica. Esto proporciona un mecanismo de respuesta retardada en el que la respuesta inmunitaria innata tiene tiempo (~48 h) para eliminar una infección antes de que la respuesta inmunitaria adaptativa mediada por células T inflamatorias entre en acción.[8]
Recambio celular de T naive
Las células T naive se mantienen dinámicamente durante toda la vida mediante la combinación de: la producción de nuevas células por el timo, la renovación proliferativa de las células existentes y la pérdida por muerte o diferenciación.[2]
Véase también
Referencias
- Eagar T.N., Miller S.D. (2019). «16:Subtipos de linfocitos cooperadores y regulación de la respuesta inflamatoria». En Rich R.R., Fleisher T.A., Shearer W.T., Schroeder H., Frew A.J., Weyand C.M., ed. Inmunología Clínica: Principios y Práctica. Elsevier Health Sciences. p. 235. Consultado el 21 de diciembre de 2020.
- Mold J.E., Réu P., Olin A., Bernard S., Michaëlsson J., Rane S., et al. (2019). «Dinámica de generación celular subyacente a la homeostasis de células T ingenuas en humanos adultos». PLoS Biol 17 (10): e3000383. doi:10.1371/journal.pbio.3000383.
- De Rosa, S. C.; Herzenberg, L.A.; Herzenberg, L. A.; Roederer, M. (2001-02). «11-color, 13-parameter flow cytometry: identification of human naive T cells by phenotype, function, and T-cell receptor diversity». Nature Medicine 7 (2): 245-248. ISSN 1078-8956. PMID 11175858. doi:10.1038/84701.
- van den Broek, Theo; Borghans, José A. M.; van Wijk, Femke (8 de marzo de 2018). «The full spectrum of human naive T cells». Nature Reviews. Immunology. ISSN 1474-1741. PMID 29520044. doi:10.1038/s41577-018-0001-y.
- Gattinoni, Luca; Lugli, Enrico; Ji, Yun; Pos, Zoltan; Paulos, Chrystal M.; Quigley, Máire F.; Almeida, Jorge R.; Gostick, Emma et al. (18 de septiembre de 2011). «A human memory T cell subset with stem cell-like properties». Nature Medicine 17 (10): 1290-1297. ISSN 1546-170X. PMC 3192229. PMID 21926977. doi:10.1038/nm.2446.
- Pulko, Vesna; Davies, John S.; Martinez, Carmine; Lanteri, Marion C.; Busch, Michael P.; Diamond, Michael S.; Knox, Kenneth; Bush, Erin C. et al. (August 2016). «Human memory T cells with a naive phenotype accumulate with aging and respond to persistent viruses». Nature Immunology 17 (8): 966-975. ISSN 1529-2916. PMC 4955715. PMID 27270402. doi:10.1038/ni.3483.
- White, Jason T.; Cross, Eric W.; Kedl, Ross M. (June 2017). «Antigen-inexperienced memory CD8+T cells: where they come from and why we need them». Nature Reviews. Immunology 17 (6): 391-400. ISSN 1474-1741. PMC 5569888. PMID 28480897. doi:10.1038/nri.2017.34.
- von Essen, Marina Rode; Kongsbak, Martin; Schjerling, Peter; Olgaard, Klaus; Ødum, Niels; Geisler, Carsten (2010-04). «Vitamin D controls T cell antigen receptor signaling and activation of human T cells». Nature Immunology (en inglés) 11 (4): 344-349. ISSN 1529-2916. doi:10.1038/ni.1851.
- Rudd, Christopher E.; Trevillyan, James M.; Dasgupta, Jai Dev; Wong, Linda L.; Schlossman, Stuart F. (1 de septiembre de 2010). «Pillars article: the CD4 receptor is complexed in detergent lysates to a protein-tyrosine kinase (pp58) from human T lymphocytes. 1988». Journal of Immunology (Baltimore, Md.: 1950) 185 (5): 2645-2649. ISSN 1550-6606. PMC 3791413. PMID 20724730.
| Control de autoridades |
|
|---|
 Datos: Q6959842
Datos: Q6959842