Calanoida
Los calanoides (Calanoida) son un orden de copépodos que se compone de especies con anténulas segmentadas entre 16 y 26 segmentos y antenas birrámeas.[2] El orden está compuesto por 42 familias con 289 géneros de las cuales solo cuatro familias están presentes en agua dulce y otras 5 están tanto en agua dulce como en agua salada.[2] Los miembros de este orden son particularmente abundantes en los ambientes pelágicos de estuarios y otros hábitats costeros.[3] Adaptados a la vida planctónica, son considerados como un grupo homogéneo por sus caracteres similares y exitoso por su gran abundancia.[3] Por lo general, representan una importante conexión entre el fitoplancton y los peces.[3] Los miembros de este orden miden en promedio 1 mm, con especies tan grandes como 10 mm y tan pequeñas como 0.4 mm.[3] Asimismo, Cuenta con 75% de especies marinas y 25% de agua salada. Algunas especies marinas son bento-pelágicas o comensales.[4]
| Calanoida | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Animalia | |
| Subreino: | Bilateria | |
| Superfilo: | Ecdysozoa | |
| Filo: | Arthropoda | |
| Subfilo: | Crustacea | |
| Clase: | Copepoda | |
| Infraclase: | Neocopepoda | |
| Superorden: | Gymnoplea | |
| Orden: |
Calanoida G.O. Sars, 1903[1] | |
Morfología externa
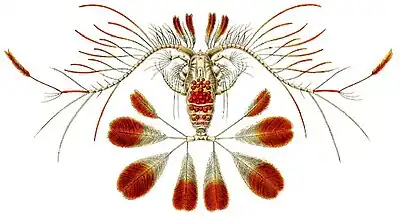
Este orden, presenta una morfología muy diversa. El cuerpo se suele dividir en cefalosoma, metasoma y urosoma. El cefalosoma se compone por la unión de la cabeza con el segmento que contiene a los maxilipedos. En cambio el metasoma esta formado por los segmentos que presentan patas y el urosoma es la parte posterior del cuerpo. En ese sentido, el prosoma se compone del cefalosoma y el metasoma.[5] El abdomen es angosto y el corazón esta presente.[2] Además, se caracterizan porque sus anténulas son más largas que su cuerpo.
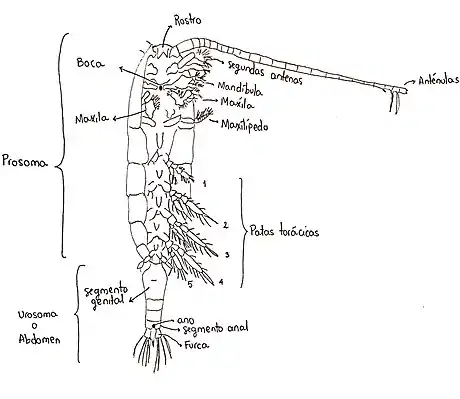
Labio y labro
El labio y el labro normalmente no son considerados con los apéndices pareados pero en conjunto forman los márgenes de la boca. El labro es un lóbulo muscular, el cual forma el margen anterior de la boca, a veces está provisto de ornamentación como espinas y contiene ocho glándulas labrales, cuatro en cada lado. Estas glándulas producen secreciones que se unen a la comida e inician la digestión en la cavidad bucal.[4]
Los lóbulos pareados del labio forman los márgenes posteriores y parte del margen lateral de la boca. Estos lóbulos pareados se derivan se la fusión de los paragnaths.[4]
Anténula
La hembra típica de este grupo posee una antena con 25 segmentos, sin embargo, la fusión de los segmentos es común en muchas especies, como Pontellopsis que muestra tan solo unos 16 segmentos. Las anténulas de hembras y muchos machos presentan simetría bilateral. Algunos machos de ciertas familias muestran una asimetría bilateral y se debe a que la anténula derecha está geniculada, eso significa que tienen una articulación a modo rodilla que separa las regiones distal y proximal. Sin embargo, machos de la superfamilia Arietelloidea tienen la anténula izquierda geniculada. Asimismo, hay géneros dentro de esa superfamilia que tienen la anténula derecha o izquierda geniculada e incluso en especies del género Pleuromamma esto varía con el individuo.[4]
Las anténulas están provistas de setae y sensilla. Por último, las anténulas del macho son usadas para el agarre de la hembra durante el apareamiento.[4]
Antena
La antena contrariamente a la anténula es un apéndice birrámeo con un endópodo y un exópodo. El endópodo usualmente consta de 3 segmentos, de los cuales el segundo y el tercero están parcialmente fusionados. El exópodo contiene de 8 a 9 segmetos y sus patrones de fusión varían. La antena en conjunción con las otras piezas bucales forma una parte integral del mecanismo de acople de comida al igual que el mecanismo de agarre del copépodo.[4]
Mandíbula
La mandíbula es birrámea, cuenta con un endópodo de 2 segmentos y un exópodo de 5 segmentos. Los segmentos basales forman la gnathobase con su espinoso y distal borde que macera la comida. El número y la forma de las espinas (dientes) al igual que la setulación de tanto el endópodo como el exópodo dependen de la dieta de los individuos.[4]
La forma de la gnathobase y la disposición de las espinas es tal que sirve para identificar especies.[4]
Maxila 1
Es un complejo apéndice laminar, es birrámeo con un endópodo de 3 segmentos, algunos a veces están fusionados y un exópodo de un solo segmento. La setulación y la forma general se relacionan con la dieta de las especies.[4]
Maxila 2
Es también un apéndice laminar, es unirrámeo y tiene 7 segmentos. Su forma se relaciona con la dieta de las especies, espinas fuertes remplazan las setas en algunas especies.[4]
Maxilípedo
Es unirrámeo y cuenta con 9 segmentos, a pesar del grueso de las especies solamente cuenta con 6 segmentos libres en su endópodo. Puede estar altamente desarrollado y armado con setas o espinas dependiendo de la estrategia alimenticia.[4]
Morfología interna
Sistema esquelético- muscular
El esqueleto de este grupo esta constituido por formaciones granulares o laminares de tipo tendinoso. La unión de los músculos con el endoesqueleto se da a partir de estructuras semejantes a los tendones. Presentan músculos longitudinales (ventrales y dorsales) que permiten conectar las dos partes principales del cuerpo: la parte anterior y la posterior. Asimismo, presentan músculos que van desde el quinto segmento torácico al segmento abdominal.[5] En cuanto al endoesqueleto, este provee el soporte para que los músculos se anclen.[4]
Aparato digestivo
Presentan dos líneas de dientes quitinosos. En su boca tienen 8 glándulas, que secretan un mucus que se mezcla con la comida y protege al intestino de la enzimas digestivas. La boca se encuentra conectada con el esófago y este con el intestino medio. El intestino esta dividido en 3 zonas: anterior, media y posterior.[5]
Sistema excretor
El sistema excretor cuenta con dos glándulas: antenulares y maxilares. Las primeras, desembocan en la base de las antenas y funcionan únicamente en la larva nauplio. Los segundos, por su parte desembocan en la base de las segundas maxilas.[5]
Sistema circulatorio
Tienen un sistema circulatorio abierto, de tipo lagunar y a diferencia de los demás copépodos, este grupo posee un corazón. Este se encuentra entre el segundo y tercer segmento torácico. Presenta cuatro aperturas, una comunica a la aorta y las otras al pericardio. El líquido circulatorio es la hemolinfa, la cual después de pasar por las vísceras, va al pericardio y por el movimiento del corazón para a la aorta. De ahí de mueve a la parte anterior y luego a la parte posterior del cuerpo. Por último, se devuelve al pericardio, donde inicia nuevamente el ciclo.[5]
Sistema respiratorio
Dado su pequeño tamaño la respiración en este grupo es cutánea. De esa forma, la respiración se da a través del tegumento.
Sistema nervioso
Como los demás Crustáceos, este grupo presenta un protocerebro que envía ramas nerviosas a los órganos frontales (ojos). Luego, se encuentra el deutocerebro, de cada lado del collar salen los nervios de las primeras antenas. Finalmente, se encuentra el tritocerebro, donde de cada lado del collar salen los nervios de las segundas antenas.[7] De igual forma, presentan un cordón ventral del cual salen las raíces dorsales que son motoras y las ventrales que son sensoriales [5]
Órganos sensoriales
Pocas familias de este grupo tienen órganos sensoriales aparentes. Algunos no presentan ojos en la parte posterior. En la larva presentan el ojo nauplio, el cual es simple y esta formado por 3 o 4 ocelos con fotorreceptores que permiten a los individuos determinar la situación del fondo o de la capa superficial del agua. Esta estructura, se va modificando desde las primeras fases del desarrollo con el objetivo de perfeccionar la fotorrecepción. Por otra parte, presentan otros órganos sensoriales como: órgano de Gicklhorn, el de Bellonci y el pleural cefálico con funciones quimiorreceptoras o fotorreceptoras. También, tienen estetes (quimiorreceptores) y sedas sensoriales (mecanorreceptores) que se encuentran en las anténulas.[5]
Aparato reproductor
Hembras y machos poseen una sola gónada situada debajo del pericardio. El macho por su parte tiene un solo ducto, mientras que la hembra tiene dos.
- Macho: en estos el segmento P5 esta modificado como un órgano copulador en machos.[2] Presentan: testículo, vaso deferente, vesícula seminal, saco espermático, espermatóforo y el conducto eyaculador.[5]
- Hembra: Presenta un ovario por encima del tubo digestivo.[5]
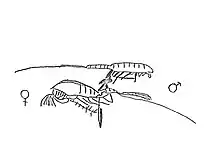
Reproducción y desarrollo
El comportamiento para el apareamiento consiste en una serie de eventos que están más o menos generalizados para todas las especies. Estos son la atracción del macho a la hembra, la captura de la hembra por parte del macho, la acomodación en la posición de apareamiento y finalmente la transferencia y la fijación del espermatóforo. El espermatóforo descarga su contenido en la espermateca de la hembra o, en las especies que no tienen espermateca se vacía dentro de una masa formada por las secreciones del espermatóforo en sí. El macho usualmente libera a la hembra inmediatamente después de la transferencia no obstante el agarre continúa en algunas especies luego del apareamiento.[8] La posición de apareamiento, el espermatóforo y el uso de la quinta pata son notables porque resuelven un problema presente en los copépodos machos, no tienen un órgano reproductor y sus espermatozoides no tienen flagelo, por lo cual la reproducción se dificulta.

Hay evidencia circunstancial que las hembras producen feromonas que atraen a los machos, que son detectadas mediante los quimiorreceptores del macho. También se ha propuesto detección mecánica de la pareja mediante mecanorreceptores, a través de la detección de las perturbaciones en el fluido producidas por la hembra. Normalmente se aproxima a la hembra por detrás y se dispone a la posición de apareamiento. Sin embargo, se han observado casos de machos aproximándose a hembras de otras especies por lo cual es posible que la quimio-mecanorrecepción inicial no siempre es efectiva. Para ello un reconocimiento químico secundario cuando el macho se prepara para la posición de apareamiento al igual que el no encaje de las piezas sexuales, se combinan para prevenir la hibridación.[8]
Muchos miembros de este orden producen huevos subitáneos, que eclosionan inmediatamente, y también huevos de diapausa que permanecen dormidos por largos periodos de tiempo.[3]
El desarrollo post-embriónico de los copépodos está dividido en dos fases, una fase nauplio y una fase de copepodito. A modo de carácter primitivo cada una de estas fases cuenta con seis etapas, usualmente llamadas NI, NII, NIII, etc y CI, CII, etc, respectivamente.[10]
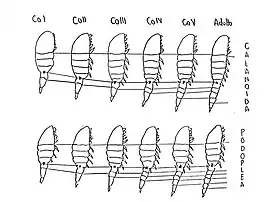
Las tres primeras fases de nauplio son verdaderos nauplios con sus tres pares de apéndices, luego las fases posteriores se parecen a metanauplios de otros crustáceos porque en general tienen signos de segmentación abdominal y rudimentos de apéndices posteriores.[4]
Los copépodos carecen de cualquier expresión de somitas durante su fase nauplio. El cuerpo naupliar generalmente crece con cada paso de una etapa a la otra. Los nauplios tempranos cuentan con tres apéndices bien desarrollados (anténulas, antena y mandíbulas) y las pequeñas yemas del rami caudal. Yemas de otros apéndices entre la mandíbula y el ramus caudal son añadidas durante la fase naupliar. Los miembros del orden Calanoida poseen las seis fases de nauplio. Por otro lado, en el estado de copepodito (durante sus fases es como un adulto en miniatura[4]) usualmente tienen las somitas abdominales y torácicas separadas por una membrana, carecen del endito naupliar en la coxa de la antena, exhiben un buen desarrollo de los apéndices post-mandíbulares y tienen sus patas nadadoras anidadas por un esclerito intercoxal. La antena se mueve de una posición paraoral en el nauplio a una pre-oral en el copepodito y pierde su función masticatoria. Durante cada muda una nueva somita es agregada. Los miembros del orden Calanoida también presentan las seis fases del estado copepodito.[10]
Alimentación
La alimentación de los copépodos está dividida en dos principales modos, alimentación por emboscada y alimentación por corrientes alimenticias. Algunos pocos copépodos del orden Calanoida usan el estilo emboscada, mientras la mayoría (pelágicos), generan corrientes alimenticias.[11] Esto se logra a través de hacer vibrar los apéndices alimentarios, y en algunas especies también usan sus patas nadadoras, en patrones regulares y complejos [11] Así con esos movimientos armónicos son capaces de crear una corriente que se dirige hacia ellos, arrastrando las presas con ella. En la mayoría de las especies la corriente alimenticia no es pasada a través de un filtro, sino que se comporta como una corriente que escanea. Es decir, las presas que vienen con la corriente alimenticia son percibidas y capturadas individualmente a través de su maxila que se ensancha en respuesta a la presa que arriba. La detección de las presas en la corriente alimenticia se debe posiblemente a pistas químicas, la nube de solutos que rodea a la presa es sacada y llevada directamente al copépodo a través de la corriente. De tal manera que la presa no tiene oportunidad para reaccionar ya que la señal química ha llegado al copépodo depredador mucho antes que a la presa en sí.[11] La concentración de la nube de solutos depende del tamaño de la presa por consiguiente las presas más grandes pueden ser detectadas a mayores distancias. Asimismo, también implica que la tasa de despeje de atrapar grandes es mayor que presas pequeñas, lo cual es consistente con las observaciones en la naturaleza donde estos tienen cierta preferencia por las presas más grandes. De igual manera, este mecanismo de detección implica un límite inferior en las presas, unos 10µm, pero esto no es consistente con las observaciones en la naturaleza porque los copépodos son capaces de atrapar presas por debajo de este límite, lo cual sugiere la existencia de otro mecanismo para esos tamaños. No obstante, ese mecanismo no ha sido claramente observado, descrito, ni entendido cuantitativamente.[11]
La alimentación por corriente alimenticia es inherentemente más eficiente que el estilo emboscada, por dos razones; el estilo emboscada está restringido a presas móviles mientras que el estilo corriente alimenticia no está limitado y funciona tanto para presas móviles como no móviles. En la corriente alimenticia la velocidad de encuentro está gobernada por la velocidad de la corriente mientras que en el estilo emboscada la velocidad de encuentro está gobernada por la velocidad a la que nada la presa. Con esta última siendo 10 veces menor, es decir se tarda 10 veces más en encontrarse con la presa.[11]
Por último, a pesar de prácticamente depender de ambos o un solo mecanismo de alimentación y vivir en la columna de agua, algunos copépodos son consistentemente hallados en agregados comiendo sobre las superficies. Observaciones y modelos han sugerido que este modo de alimentarse puede resultar cuantitativamente importante.
Hábitat y distribución
En este orden, la mayoría son pelágicos llegando a habitar aguas saladas, dulces en capas de aguas superficiales o profundas. En cuanto a los demás, suelen ser bentónicos, es decir que habitan sobre el sedimento o en los primero milímetros de este. Además, algunos pueden ser parásitos de peces, localizados en las branquias.[5]
Tienen una distribución cosmopolita; se encuentran en todos los océanos y mares del mundo, así como en lagos, lagunas y charcascharcas. Asimismo, es importante destacar que dependiendo de las características de la masa de agua diferentes poblaciones dominan ese medio.[5]
Filogenia
Ubicación en la clase Copepoda
Esta filogenia se basa en el análisis molecular las subunidades ribosomales 18s y 28s, proteína histona H3 y COI mtDNA. A pesar de su abundancia, las relaciones entre los órdenes y la monofilia de algunos de ellos no estaban bien definidos, hasta la realización de este estudio.[12]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||







Clasificación interna
Debido a la complejidad morfológica en este grupo la subdivisión y filogenia han sido complejas y problemáticas, a diferencia del orden, que ha sido taxonómicamente bien definido. Esta filogenia esta basada en análisis molecular del 18s y el 28s, que son subunidades ribosomales.[13]
| Calanoida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Clasificación
La clasificación de este orden es la siguiente:[14]
- Familia Acartiidae
- Familia Aetideidae
- Familia Arietillidae
- Familia Augaptilidae
- Familia Bathypontiidae
- Familia Boholinidae
- Familia Calanidae
- Familia Candaciidae
- Familia Centropagidae
- Familia Clausocalanidae
- Familia Diaixidae
- Familia Diaptomidae
- Familia Discoidae
- Familia Epacteriscidae
- Familia Eucalanidae
- Familia Euchaetidae
- Familia Fosshageniidae
- Familia Heterohabdidae
- Familia Hyperbionychidae
- Familia Lucicutiidae
- Familia Mecynoceridae
- Familia Megacalanidae
- Familia Mesaiokeratidae
- Familia Metridinae
- Familia Nullosetigeridae
- Familia Paracalanidae
- Familia Parapontellidae
- Familia Parkiidae
- Familia Phaenidae
- Familia Pseudocyclopidae
- Familia Pseudocyclopiidae
- Familia Pseudodiaptomidae
- Familia Rhincalanidae
- Familia Ridgewayiidae
- Familia Ryocalanidae
- Familia Scolecitrichidae
- Familia Spinocalanidae
- Familia Stephidae
- Familia Sulcanidae
- Familia Temoridae
- Familia Tharybidae
- Familia Tortanidae
Véase también
Referencias
- «Calanoida». 2020.
- Bledzki, Leszek A.; Rybak, Jan Igor (2016). Freshwater Crustacean Zooplankton of Europe (en inglés británico). doi:10.1007/978-3-319-29871-9. Consultado el 23 de mayo de 2020.
- Lee, Cheng-Sheng, ed. (27 de abril de 2005). «Copepods in Aquaculture». Wiley Online Library (en inglés). doi:10.1002/9780470277522. Consultado el 23 de mayo de 2020.
- «The Biology of Calanoid Copepods, Volume 33 - 1st Edition». www.elsevier.com. Consultado el 24 de mayo de 2020.
- Fernández, Maria Luz. «Orden Calanoida». 2015 (89): 1-27.
- Fernández, Maria Luz. «Orden Calanoida». 2015.
- Mooi, Rich; Brusca, Richard C.; Brusca, Gary J. (1991-06). «Invertebrates.». Systematic Zoology 40 (2): 245. doi:10.2307/2992263. Consultado el 24 de mayo de 2020.
- «The Biology of Calanoid Copepods, Volume 33 - 1st Edition». www.elsevier.com. Consultado el 24 de mayo de 2020.
- «Atlas of Crustacean Larvae | Johns Hopkins University Press Books». jhupbooks.press.jhu.edu. Consultado el 24 de mayo de 2020.
- «Atlas of Crustacean Larvae | Johns Hopkins University Press Books». jhupbooks.press.jhu.edu. Consultado el 24 de mayo de 2020.
- Kiørboe, Thomas (2011-05). «What makes pelagic copepods so successful?». Journal of Plankton Research (en inglés) 33 (5): 677-685. ISSN 1464-3774. doi:10.1093/plankt/fbq159. Consultado el 24 de mayo de 2020.
- Khodami, Sahar; McArthur, J. Vaun; Blanco-Bercial, Leocadio; Martinez Arbizu, Pedro (22 de agosto de 2017). «Molecular Phylogeny and Revision of Copepod Orders (Crustacea: Copepoda)». Scientific Reports (en inglés) 7 (1): 1-11. ISSN 2045-2322. doi:10.1038/s41598-017-06656-4. Consultado el 24 de mayo de 2020.
- Blanco-Bercial, Leocadio; Bradford-Grieve, Janet; Bucklin, Ann (1 de abril de 2011). «Molecular phylogeny of the Calanoida (Crustacea: Copepoda)». Molecular Phylogenetics and Evolution (en inglés) 59 (1): 103-113. ISSN 1055-7903. doi:10.1016/j.ympev.2011.01.008. Consultado el 24 de mayo de 2020.
- «ITIS Standard Report Page: Calanoida». www.itis.gov. Consultado el 23 de mayo de 2020.
| Control de autoridades |
|
|---|