Conmutación de clase de inmunoglobulina
La conmutación de clase de inmunoglobulina (o conmutación del isotipo o conmutación isotípica o recombinación del cambio de clase) es un mecanismo biológico que hace cambiar al anticuerpo de una clase a otra, por ejemplo, del isotipo conocido como IgM al isotipo llamado IgG. Durante este proceso se cambia la porción de región constante de la cadena pesada del anticuerpo, pero la región variable permanece sin cambios. Puesto que ésta no cambia, esta conmutación no afecta a la especificidad de antígeno. En lugar de ello, el anticuerpo conserva su afinidad por los mismos antígenos, pudiendo sin embargo interactuar con diferentes moléculas efectoras.
Dominique Butrón (2017:): "El class switching es un proceso en el que se cambia el anticuerpo de una clase por el anticuerpo de otra. En este proceso se cambia la porción constante de la cadena pesada, mientras que se mantiene la porción variable. La porción constante sirve para unión a las células efectoras, y la porción variable permite la unión al epitope del antígeno, por lo que el anticuerpo puede unirse a más células efectoras, sin cambiar su afinidad por el antígeno".
Mecanismo
La conmutación de la clase tiene lugar tras la activación de un linfocito B maduro a través de su anticuerpo unido a membrana (también conocido como receptor del linfocito B) para generar los distintos tipos de anticuerpo, todos ellos con los mismos dominios variables que el anticuerpo original generado en el linfocito B inmaduro durante el proceso de recombinación V(D)J, aunque con distintos dominios constantes en su cadena pesada.[1]
Los linfocitos B vírgenes producen tanto IgM como IgD, que son los dos primeros segmentos de la cadena pesada en el locus de la inmunoglobulina. Tras su activación por antígeno, estos linfocitos B proliferan y comienzan a producir altos niveles de anticuerpos. Si estos linfocitos B se activan por sus receptores de CD40 y de IL-4 (ambos modulados por los linfocitos T colaboradores), sufren la conmutación de isotipo para producir anticuerpos IgG, IgA o IgE. Al permanecer constante la región variable de las cadenas pesadas, se producen diferentes progenies a partir del mismo linfocito B activado con distintos isotipos o incluso subtipos (p.ej. IgG1, IgG2, etc.).[2]
El orden de los diferentes exones de la cadena pesada es el siguiente:
- μ - IgM
- δ - IgD
- γ3 - IgG3
- γ1 - IgG1
- pseudogén similar al gen ε que ha perdido la función.
- α1 - IgA1
- γ2 - IgG2
- γ4 - IgG4
- ε - IgE
- α2 - IgA2
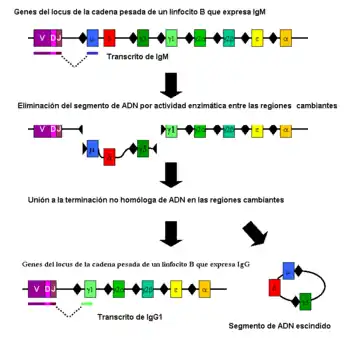
La conmutación de clase tiene lugar mediante un mecanismo conocido como unión tras la recombinación en el cambio de clase "class switch recombination" en inglés (CSR). Durante este proceso, porciones del locus de la cadena pesada del anticuerpo son eliminadas del cromosoma y los segmentos génicos que rodean la porción destruida se empalman para mantener un gen funcional de anticuerpo, pero que produce un isotipo diferente. Se generan rupturas en ambas cadenas del ADN en motivos de nucleótidos conservados, llamados regiones (s), que se encuentran "corriente arriba" (son anteriores en la secuencia de ADN) de los segmentos génicos que codifican las regiones constantes de las cadenas pesadas del anticuerpo; esto tiene lugar en un punto adyacente a todos los genes de las regiones constantes de las cadenas pesadas con la excepción de la cadena δ. El ADN sufre una melladura (Nick) y se rompe en dos regiones S seleccionadas por la actividad de una serie de enzimas, como la citidina deaminasa inducida por activación(AID), uracil ADN glicosilasa específica de uracilo y la endonucleasa apurínica/apirimidínica (endonucleasas AP).[3][4] El ADN situado entre las regiones S es, de ese modo, eliminado del cromosoma, retirando las regiones correspondientes a las cadenas pesadas μ o δ y sustituyéndolas por un segmento de cadena pesada γ, α o ε. Los extremos libres del ADN se reúnen por un proceso denominado unión de extremos no homóloga (non-homologous end joining, NHEJ) para ensamblar el exón del dominio variable con el exón de cadena constante codificado "corriente abajo".[5] En ausencia de este proceso, los extremos libres de ADN se pueden reunir por una ruta alternativa derivada hacia uniones de microhomología.[6] Con excepción de los genes μ y δ solo se expresa una clase de anticuerpo cada vez en cualquier momento.
Véase también
- Immunogenética
Referencias
- Eleonora Market, F. Nina Papavasiliou (2003) V(D)J Recombination and the Evolution of the Adaptive Immune System PLoS Biology1(1): e16.
- Stavnezer J, Amemiya CT (2004). «Evolution of isotype switching». Semin. Immunol. 16 (4): 257-75. PMID 15522624. doi:10.1016/j.smim.2004.08.005.
- Durandy A (2003). «Activation-induced cytidine deaminase: a dual role in class-switch recombination and somatic hypermutation». Eur. J. Immunol. 33 (8): 2069-73. PMID 12884279. doi:10.1002/eji.200324133.
- Casali P, Zan H (2004). «Class switching and Myc translocation: how does DNA break?». Nat. Immunol. 5 (11): 1101-3. PMID 15496946. doi:10.1038/ni1104-1101.
- Lieber MR, Yu K, Raghavan SC (2006). «Roles of nonhomologous DNA end joining, V(D)J recombination, and class switch recombination in chromosomal translocations». DNA Repair (Amst.) 5 (9-10): 1234-45. PMID 16793349. doi:10.1016/j.dnarep.2006.05.013.
- Yan CT, Boboila C, Souza EK, Franco S, Hickernell TR, Murphy M, Gumaste S, Geyer M, Zarrin AA, Manis JP, Rajewsky K, Alt FW (2007). «IgH class switching and translocations use a robust non-classical end-joining pathway». Nature 449: 478-82. PMID 17713479. doi:10.1038/nature06020.