Cristatusaurus lapparenti
Cristatusaurus lapparenti ("repil crestado de Lapparent") es la única especie conocida del género extinto Cristatusaurus de dinosaurio terópodo espinosáurido, que vivió a mediados del período Cretácico, hace aproximadamente 100 millones de años, en el Albiense, en lo que es hoy África. Solo se conoce una parte de la mandíbula de 22 centímetros y un diente, con esto se ha propuesto que llegó a medir 10 metros de largo, 4 de alto y a pesar 1,5 tonelada.
| Cristatusaurus lapparenti | ||
|---|---|---|
| Rango temporal: 100 Ma Cretácico inferior | ||
 Cristatusaurus lapparenti, mandíbula fósil, Museo Nacional de Historia Natural de Francia, París | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| (sin rango): | Tetanurae | |
| Superfamilia: | Megalosauroidea | |
| Familia: | Spinosauridae | |
| Subfamilia: | Baryonychinae | |
| Género: |
Cristatusaurus Taquet & Russell, 1998 | |
| Especie: |
C. lapparenti Taquet & Russell, 1998 | |
Existe cierto debate sobre la identidad de esta criatura, en la que algunos argumentan que probablemente sea el mismo dinosaurio que Suchomimus, que también se ha encontrado en Níger, en las mismas capas. En ese caso, el nombre Cristatusaurus tendría prioridad. Otros han concluido, sin embargo, que Cristatusaurus es un nomen dubium , ya que es indistinguible tanto de Suchomimus como de Baryonyx.[1][2] Mientras tantos otros autores lo consideran como un Barionyx[3][4]
Descripción
En 2012, el paleontólogo vertebrado estadounidense Thomas R. Holtz Jr. calculó tentativamente el Cristatusaurus en alrededor de 10 metros de largo y un peso de 1 a 4 toneladas.[5][6] Los premaxilares de holotipo son de 115 milímetros de largo y 55 milímetros de alto. El otro conjunto conocido de premaxilares, la muestra MNHN GDF 365 es más grande con 185 milímetros de largo y 95 milímetros de alto.[7] El tamaño más pequeño del holotipo, la superficie más lisa y la falta de suturas osificadas indican que pertenece a un individuo juvenil, mientras que MNHN GDF 365 probablemente representa a un adulto.[7]

La punta del premaxilar de Cristatusaurus fue corta y expandido, mientras que el extremo trasero se redujo cerca de la sutura con el maxilar superior. Esta forma de hocico como roseta era característica de los espinosáuridos. La parte frontal de la mandíbula superior era cóncava en la parte inferior, con una forma que se entrelazaba con lo que habría sido la punta convexa y también agrandada del hueso dentario de la mandíbula.[8][7][9] Una delgada cresta sagital corría longitudinalmente sobre los premaxilares, una condición presente en Baryonyx y Suchomimus y muy prominente en Angaturama, un posible sinónimo de Irritator.[10] Como todos los espinosáuridos, las narinas externas de Cristatusaurus ' s narinas fueron posicionados más atrás en el cráneo que en terópodos típicos.[7][10] Dos procesos óseos se extienden por la parte inferior del hocico, en una estructura convexa que forma el paladar secundario del animal. Esta condición se observa en todos los cocodrilos actuales, pero no en la mayoría de los dinosaurios terópodos; sin embargo, era un rasgo común entre los espinosáuridos.[10][11]
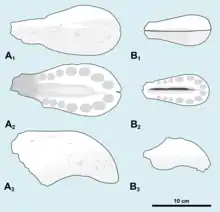
Los alvéolos dentales de Cristatusaurus estaban estrechamente espaciados, en el maxilar y dentario se aplanaron algo de lado, mientras que los de los premaxilares eran grandes y en su mayoría circulares, siendo los más grandes los alvéolos frontales.[8][7] Las coronas de dientes parciales conservadas en algunos alvéolos muestran que los dientes estaban serrados finamente, con estrías en su superficie lingual, el lado de los dientes que mira hacia adentro.[7] Ambas muestras de premaxilares tenían siete alveolos en cada lado, el mismo número que en Suchomimus , Angaturama , Oxalaia y el epécimen de Spinosaurus maroccanus.[8][12]
Una de las vértebras dorsales asignadas, MNHN GDF 358 medía 13,5 centímetros de longitud central, que es igual a la vértebra más grande conocida de Spinosaurus maroccanus. La base conservado de una espina neural vertebral de Cristatusaurus, MNHN GDF 359 mide 15 milímetros de espesor en comparación con el 25 milímetros visto en una equivalente de Spinosaurus lo que indica que las espinas neurales de Cristatusaurus probablemente no eran tan altas como los de Spinosaurus.[8] De los dos unguales manuales en forma de garras referidos a Cristatusaurus, uno era equivalente en tamaño a los encontrados para Suchomimus y Baryonyx , mientras que el otro era aproximadamente 25 a 30 por ciento más pequeño.[13] Como un espinosáurido, habría manejado estas garras con manos de tres dedos llevadas por brazos robustos.[9]
Descubrimiento e investigación
Sus fósiles fueron encontrados por Philippe Taquet en Gadoufaoua en Níger en 1973. Estos fósiles son bastante similares a los de Baryonyx y Suchomimus. La especie tipo , Cristatusaurus lapparenti, fue descrita formalmente por Taquet y Dale Russell en 1998. El nombre genérico se refiere a una cresta, crista en latín , en el hocico. El nombre específico honra al fallecido Albert-Félix de Lapparent.[8] El holotipo, MNHN GDF 366, fue encontrado en las camas Tegema que datan de finales del Aptiano. Consiste en dos premaxilares desarticuladas , un maxilar derecho y un fragmento de mandíbula inferior, de un individuo juvenil.[7] Se han asignado varios paratipos, MNHN GDF 365, un hocico de dos praemaxilares articuladas y MNHN GDF 357, 358, 359 y 361, cuatro vértebras dorsales.[8][14]

Los primeros fósiles de Cristatusaurus fueron encontrados en 1973 por el paleontólogo francés Philippe Taquet en Gadoufaoua, una localidad dentro de la formación Elrhaz en Níger. El espécimen holotipo , catalogado con el número MNHN GDF 366, consta de dos premaxilares, huesos del hocico frontal, un maxilar parcial derecho, el hueso de la mandíbula superior principal y un fragmento de la dentadura de la mandíbula . Varios paratipos han sido asignados, MNHN GDF 365, un hocico de dos premaxilares articulados, así como MNHN GDF 357, MNHN GDF 358, MNHN GDF 359 y MNHN GDF 361, cuatro vértebras dorsales.[15][8] Dos garras de pulgar de especímenes separados también se atribuyeron posteriormente a Cristatusaurus.[13] En 1984, los especímenes de premaxilares MNHN GDF 365 y MNHN GDF 366¡ fueron descritos por primera vez por Taquet, donde los remitió a un nuevo terópodo sin nombre dentro de la familia Spinosauridae, debido a sus características compartidas con el holotipo dentario de Spinosaurus aegyptiacus.[15] En el momento en que Taquet creía que estos especímenes pertenecían a la mandíbula inferior de la criatura, ya que no se conocía ningún terópodo con más de cinco dientes en la premaxilar, mientras que Cristatusaurus tenía siete. Esto se demostró posteriormente como incorrecto en 1996 por los paleontólogos brasileños Alexander Kellner y Diógenes Campos, a la luz de los descubrimientos de otros espinosáuridos que conservan las puntas de la mandíbula superior con más de cinco dientes.[15][7]
En una publicación de 1986, los paleontólogos británicos Alan Charig y Angela Milner consideraron los elementos de la mandíbula de Taquet casi indistinguibles de los del espinosáurido Baryonyx walkeri, que describían sobre la base de un esqueleto parcial del Barremiano de la formación de Arcilla Weald, Inglaterra.[16] Una continuación de 1997 de este documento preliminar remitió a MNHN GDF 365 y MNHN GDF 366 a una especie de Baryonyx indeterminada , independientemente de su edad geológica más joven.[3] En 1998 , Taquet y el geólogo estadounidense Dale Russell utilizaron los huesos para erigir el nuevo género Cristatusaurus, con la especies de tipo siendo Cristatusaurus lapparenti . Su nombre genérico se deriva del latín crista, por "cresta" y se refiere a una cresta sagital en el hocico.[8] El nombre específico honra al fallecido paleontólogo francés Albert-Félix de Lapparent, debido a sus contribuciones a los descubrimientos relacionados con los dinosaurios en el Sahara. En el mismo artículo, varios cráneos y fósiles vertebrales del Tademait de Argelia fueron atribuidos a una nueva especie de Spinosaurus llamado S. maroccanus, que fue descrito y comparado con Cristatusaurus.[8] Spinosaurus maroccanus ahora es considerado por la mayoría de los paleontólogos como un dudoso[17][4][18] o uno sinónimo de S. aegyptiacus.[2] Dos meses después de que Taquet y Russel publicaran su artículo, otro género y especie espinosáurido fue nombrado de la formación Erlhaz, Suchomimus tenerensis. Sus descriptores, el paleontólogo estadounidense Paul Sereno y sus colegas, estuvieron de acuerdo con Charig y Milner en que no había distinción entre los fósiles de cráneo de Baryonyx y Cristatusaurus y concluyendo que este último era un nomen dubium.[2] En un análisis de 2003, el paleontólogo alemán Oliver Rauhut estuvo de acuerdo con esto.[18]
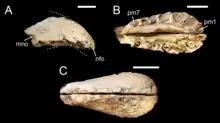
Al describir el taxón , Taquet y Russel basan la separación de Cristatusaurus de Baryonyx en la condición brevirostra, de hocico corto, del premaxilar.[8] El significado de este diagnóstico ha sido considerado oscuro por varios autores posteriores, que describen los especímenes como casi idénticos a los de Baryonyx y Suchomimus.[19] En 2002, Eric Buffetaut y Mohamed Ouaja apoyaron la sinonimia de Cristatusaurus con Baryonyx.[4] El mismo año, Hans-Dieter Sues y sus colegas consideraron tanto a Cristatusaurus como a Suchomimus como sinónimos menores de Baryonyx, afirmando que no hay evidencia fósil que indique que más de un espinosáurido vivió en la formación Elrhaz.[11] Investigaciones más recientes han conservado Suchomimus y Baryonyx como géneros distintos.[10][20][21] Otros, como Bertin Tor en 2010, y Carrano y sus colegas en 2012, se han referido a Cristatusaurus como una barioniquino indeterminado, debido a lo fragmentarios que son sus restos.[19][22]
En 2016, Christophe Hendrickx, Octávio Mateus y Buffetaut notaron que Taquet y Russel podrían haber interpretado que Cristatusaurus tenía un hocico más corto que Baryonyx al confundir la muesca donde el maxilar se articulaba con el premaxilar con las aberturas de la ventana de la nariz. Dado que tanto Suchomimus y Baryonyx han conservado más completo premaxilares, mientras Cristatusaurus sólo tiene la parte más delantera de este hueso conocido, Hendrickx y sus colegas consideraron que era posible que el hocico de Cristatusaurus será tan largo como en Baryonyx. Por lo tanto, coincidieron con autores anteriores en la ambigüedad del diagnóstico de Taquet y Russel. Hendrickx y sus colegas declararon que dado que Cristatusaurus y Suchomimus son casi idénticos y ambos provienen de la misma unidad estratigráfica, casi con seguridad son sinónimos. Los investigadores encontraron que Cristatusaurus y Suchomimus eran similares en cuanto a que ambos tenían crestas premaxilares, relación de tamaño similar de las cavidades dentales y depresiones poco profundas frente a las aberturas de sus fosas nasales. Sin embargo, dado que estas características son menores y pueden variar dentro de las especies, así como dependiendo de la edad y el sexo, Hendrickx y sus colegas no identificaron ninguna autapomorfia definitiva en el holotipo de Cristatusaurus y por lo tanto considerado el taxón un dudoso hasta que sus restos poscraneales se examinaron más de cerca.[23] Dado que fue nombrado primero, Cristatusaurus lapparenti tiene prioridad sobre Suchomimus tenerensis en el caso de que se conviertan en sinónimos.[24]
En un estudio de 2017, Marcos Sales y Cesar Schultz compararon el holotipo de Cristatusaurus, MNHN GDF 366, con el hocico referido de Suchomimus, MNN GDF501. Ambos exhiben un borde estrecho en la parte superior de sus premaxilares. Sin embargo, el paladar secundario de Cristatusaurus es convexo y claramente visible en la vista lateral, situada bajo los dientes premaxilares, mientras que en Suchomimus es discernible sólo a través de grietas en el hocico fósil. También se señaló que si se conoce, el proceso de pendiente ascendente del maxilar de Cristatusaurus es más estrecho que en Suchomimus . Los investigadores concluyeron que se necesitan más estudios para determinar si estas diferencias son posibles autapomorfías entre los taxones, o si son el resultado de cambios ontogenéticos, de desarroll), dado que el holotipo Cristatusaurus representa a un individuo más joven.[10]
Clasificación
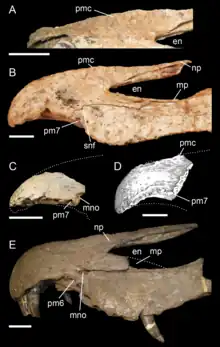
Los espinosáuridos eran grandes carnívoros bípedos con patas delanteras bien construidas y cráneos alargados con forma de cocodrilo. Las afinidades taxonómicas y filogenéticas del grupo están sujetas a investigación y debate activos, dado que en comparación con otros grupos de terópodos, muchos de los taxones de la familia, incluido Cristatusaurus se basan en material fósil relativamente pobre.[9] Tradicionalmente, la familia se ha dividido en dos subfamilias, Spinosaurinae, que incluye géneros como Irritator, Spinosaurus y Oxalaia y Baryonychinae, que incluye Baryonyx y Suchomimus. Aunque la ubicación de los géneros y especies de Cristatusaurus lapparenti está en disputa, sus fósiles pertenecen ciertamente a un miembro de la barioniquinos, debido a sus fosas nasales externas más adelantadas, relativamente primeros dientes premaxilares y las cavidades de los dientes más espaciadas que en los espinosaurinos, así como la presencia de estrías finas, en contraste con las espinosaurinos que carecen por completo de ellas.[9][11][12] Sin embargo, autores como Sales y Schultz han cuestionado la monofilia de Baryonychinae, lo que significa que podría ser un grupo no natural, afirmando que los espinosáuridos sudamericanos Angaturama e Irritator representan formas intermedias entre Baryonychinae y Spinosaurinae, en función de sus características craneales y dientes.[10]
Paleoecología
La formación Elrhaz, parte del grupo Tegama, consiste principalmente en areniscas fluviales con bajo relieve, muchas de las cuales están ocultas por las dunas de arena.[25][26] Los sedimentos son de grano grueso a medio, casi sin horizontes de grano fino.[27] Cristatusaurus vivió en lo que hoy es Níger, desde la última etapa de Aptiano hasta las primeras etapas de Albiano en el Cretácico temprano, hace 112 millones de años.[28][29] Las capas de sedimento de la formación se han interpretado como un hábitat interior de agua dulce, extensa llanuras aluviales y ríos de rápido movimiento, con un clima tropical que probablemente experimentó períodos estacionales secos.[28]

Este entorno fue el hogar de una gran variedad de fauna que incluye dinosaurios, pterosaurios, quelonios, peces, tiburones hybodontinos y bivalvos de agua dulce.[29][26] Además de Cristatusaurus lapparenti y Suchomimus tenerensis, se han encontrado terópodos como el abelisáurido Kryptops palaios , el carcarodontosáurido Eocarcharia dinops y un noasáurido desconocido. Los dinosaurios herbívoros de la región incluían iguanodontianos como Ouranosaurus nigeriensis, Elrhazosaurus nigeriensis, Lurdusaurus arenatus y dos saurópodos, Nigersaurus taqueti y un titanosauriano sin nombre. Los cocodrilomorfos eran abundantes, representado por la especie gigante de folidosáurido Sarcosuchus imperator, así como pequeños notosuquianos como Anatosuchus minor, Araripesuchus wegeneri y Stolokrosuchus lapparenti.[26] La flora local probablemente consistió principalmente de helechos, colas de caballo y las angiospermas, basadas en las adaptaciones dietéticas de los saurópodos que vivían allí.[28]
Se ha propuesto un estilo de vida semiacuático para muchos espinosáuridos, debido a sus rasgos anatómicos inusuales e histología ósea. Los dientes de Cristatusaurus probablemente se habrían usado para perforar y agarrar las presa, en lugar de cortar la carne, como lo indica su sección transversal subcircular y reducciones de estrías. Sus dientes, combinados con la curvatura similar a una onda de las mandíbulas se habría desempeñado como una trampa eficiente para los peces. Las fosas nasales retraídas le habrían permitido sumergir su hocico más bajo el agua que la mayoría de los terópodos, al tiempo que aún podían respirar y el paladar secundario óseo se ha teorizado para haber reforzado el cráneo contra las tensiones de flexión durante la alimentación. El uso de los unguales manuales recurvados gigantes de espinosáuridos todavía está en debate. Las funciones sugeridas han variado desde la captura de presas acuáticas fuera del agua, hasta la limpieza de cadáveres o la excavación.[7][11][10][30]
Referencias
- Rauhut, O.W.M. (2003). "The interrelationships and evolution of basal theropod dinosaurs". Special Papers in Palaeontology 69: 1-213.
- Sereno, P. C.; Beck, A. L.; Dutheuil, D. B.; Gado, B.; Larsson, H. C.; Lyon, G. H.; Marcot, J. D.; Rauhut, O. W. M.; Sadleir, R. W.; Sidor, C. A.; Varricchio, D. J.; Wilson, G. P.; Wilson, J. A. (1998). «A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids». Science 282 (5392): 1298-1302. Bibcode:1998Sci...282.1298S. PMID 9812890. doi:10.1126/science.282.5392.1298.
- Charig, A. J.; Milner, A. C. (1997). «Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey». Bulletin of the Natural History Museum of London 53: 11-70.
- Buffetaut, E.; Ouaja, M. (2002). «A new specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae». Bulletin de la Société Géologique de France (en inglés) 173 (5): 415-421. doi:10.2113/173.5.415. Archivado desde el original el 22 de enero de 2015. Consultado el 30 de mayo de 2019.
- Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- Holtz, T. R. Jr. (2014). «Supplementary Information to Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages». University of Maryland. Consultado el 5 de septiembre de 2014.
- Kellner, Alexander; Campos, Diogenes (1996). «First Early Cretaceous theropod dinosaur from Brazil with comments on Spinosauridae». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 199 (2): 151-166. doi:10.1127/njgpa/199/1996/151.
- Taquet, Philippe; Russell, Dale A (1998). «New data on spinosaurid dinosaurs from the early cretaceous of the Sahara». Comptes Rendus de l'Académie des Sciences, Série IIA (en inglés) 327 (5): 347-353. ISSN 1251-8050. doi:10.1016/S1251-8050(98)80054-2.
- Hone, David William Elliott; Holtz, Thomas Richard (2017). «A Century of Spinosaurs – A Review and Revision of the Spinosauridae with Comments on Their Ecology». Acta Geologica Sinica – English Edition (en inglés) 91 (3): 1120-1132. ISSN 1000-9515. doi:10.1111/1755-6724.13328.
- Sales, Marcos A. F.; Schultz, Cesar L. (2017). «Spinosaur taxonomy and evolution of craniodental features: Evidence from Brazil». PLOS ONE (en inglés) 12 (11): e0187070. Bibcode:2017PLoSO..1287070S. ISSN 1932-6203. PMC 5673194. PMID 29107966. doi:10.1371/journal.pone.0187070.
- Sues, Hans-Dieter; Frey, Eberhard; Martill, David; Scott, Diane (2002). «Irritator challengeri, a Spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil». Journal of Vertebrate Paleontology 22 (3): 535-547. doi:10.1671/0272-4634(2002)022[0535:ICASDT]2.0.CO;2.
- Kellner, Alexander W. A.; Azevedo, Sergio A. K.; Machado, Elaine B.; Carvalho, Luciana B.; Henriques, Deise D. R. (2011). «A new dinosaur (Theropoda, Spinosauridae) from the Cretaceous (Cenomanian) Alcântara Formation, Cajual Island, Brazil». Anais da Academia Brasileira de Ciências 83 (1): 99-108. ISSN 0001-3765. doi:10.1590/S0001-37652011000100006.
- «RE: JP3-Spinosaurus». dml.cmnh.org. Archivado desde el original el 13 de octubre de 2020. Consultado el 18 de septiembre de 2018.
- Machado, E. B. & Kellner, W. A. 2005. Notas sobre Spinosauridae (Theropoda, Dinosauria). Anuário do Instituto de Geociências, 28-1, 158-173.
- Taquet, Philippe (1984). «Une curieuse spécialisation du crâne de certains Dinosaures carnivores du Crétacé: le museau long et étroit des Spinosauridés.». CRAcad Sci 299: 217-222.
- Charig, Alan J.; Milner, Angela C. (1986). «Baryonyx, a remarkable new theropod dinosaur». Nature (en inglés) 324 (6095): 359-361. ISSN 0028-0836. PMID 3785404. doi:10.1038/324359a0.
- dal Sasso, C.; Maganuco, S.; Buffetaut, E.; Mendez, M. A. (2005). «New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities». Journal of Vertebrate Paleontology (Submitted manuscript) 25 (4): 888-896. ISSN 0272-4634. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2.
- Rauhut, O. W. M. (2003). «The interrelationships and evolution of basal theropod dinosaurs». Special Papers in Palaeontology 69. pp. 35-36. ISBN 978-0-901702-79-1.
- Bertin, Tor (2010). «A catalogue of material and review of the Spinosauridae». PalArch's Journal of Vertebrate Palaeontology 7.
- Benson, R. B. J.; Carrano, M. T.; Brusatte, S. L. (2009). «A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic». Naturwissenschaften (Submitted manuscript) 97 (1): 71-78. Bibcode:2010NW.....97...71B. PMID 19826771. doi:10.1007/s00114-009-0614-x.
- Carrano, Matthew T.; Benson, Roger B. J.; Sampson, Scott D. (2012). «The phylogeny of Tetanurae (Dinosauria: Theropoda)». Journal of Systematic Palaeontology (en inglés) 10 (2): 211-300. ISSN 1477-2019. doi:10.1080/14772019.2011.630927.
- Hendrickx, Christophe; Mateus, Octávio; Buffetaut, Eric (6 de enero de 2016). «Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa.». PLoS ONE 11 (1): e0144695. ISSN 1932-6203. PMC 4703214. PMID 26734729. doi:10.1371/journal.pone.0144695.
- Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (2004). The Dinosauria (en inglés). University of California Press. p. 98. ISBN 9780520941434.
- Sereno, P. C.; Beck, A. L.; Dutheil, D. B.; Larsson, H. C.; Lyon, G. H.; Moussa, B.; Sadleir, R. W.; Sidor, C. A.; Varricchio, D. J.; Wilson, G. P.; Wilson, J. A. (1999). «Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs». Science 286 (5443): 1342-1347. PMID 10558986. doi:10.1126/science.286.5443.1342.
- Sereno, P. C.; Brusatte, S. L. (2008). «Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger». Acta Palaeontologica Polonica 53 (1): 15-46. doi:10.4202/app.2008.0102.
- Sereno, P. C.; Wilson, J. A.; Witmer, L. M.; Whitlock, J. A.; Maga, A.; Ide, O.; Rowe, T. A. (2007). «Structural extremes in a Cretaceous dinosaur». PLoS ONE 2 (11): e1230. Bibcode:2007PLoSO...2.1230S. PMC 2077925. PMID 18030355. doi:10.1371/journal.pone.0001230.
- Sereno, P. C.; Wilson, J. A.; Witmer, L. M.; Whitlock, J. A.; Maga, A.; Ide, O.; Rowe, T. A. (2007). «Structural extremes in a Cretaceous dinosaur». PLoS ONE 2 (11): e1230. Bibcode:2007PLoSO...2.1230S. PMC 2077925. PMID 18030355. doi:10.1371/journal.pone.0001230..
- Sereno, Paul C.; Larson, Hans C. E.; Sidor, Christian A.; Gado, Boubé (2001). «The Giant Crocodyliform Sarcosuchus from the Cretaceous of Africa». Science 294 (5546): 1516-1519. Bibcode:2001Sci...294.1516S. PMID 11679634. doi:10.1126/science.1066521.
- Milner, Andrew; Kirkland, James (September 2007). «The case for fishing dinosaurs at the St. George Dinosaur Discovery Site at Johnson Farm». Utah Geological Survey Notes 39: 1-3.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Cristatusaurus lapparenti.
Wikimedia Commons alberga una categoría multimedia sobre Cristatusaurus lapparenti. Wikispecies tiene un artículo sobre Cristatusaurus lapparenti.
Wikispecies tiene un artículo sobre Cristatusaurus lapparenti.- Cristatusaurus en Dino directory (en inglés) Archivado el 4 de agosto de 2007 en Wayback Machine.
 Datos: Q3697359
Datos: Q3697359