Cromosoma homólogo
Los cromosomas homólogos son un par de cromosomas compuesto por un cromosoma paterno y uno materno que se emparejan dentro de una célula durante la meiosis, la cual ocurre en la reproducción sexual. Tienen la misma disposición de la secuencia de ADN de un extremo a otro, pero distintos alelos.[1] Estas copias de cromosoma tienen los mismos genes en los mismos loci donde aportan puntas a lo largo de cada cromosoma la cual facilita un par de cromosomas a alinear correctamente una con otra antes de que se separen durante la meiosis.[2][3] Ésta es la base de la herencia mendeliana, que caracteriza los patrones de herencia del material genético de un organismo a su célula de desarrollo progenitora descendiente en un momento y área determinados.[4] Existen dentro de los organismos eucariotas diploides. Las parejas de cromosomas que son diferentes se llaman heterólogos y corresponden casi siempre a los cromosomas sexuales.[5]

Resumen
Los cromosomas son disposiciones lineales de ácido desoxirribonucleico (ADN) condensado y proteínas histonas, que forman un complejo denominado cromatina.[4] Los cromosomas homólogos están formados por pares de cromosomas de aproximadamente la misma longitud, posición del centrómero y patrón de tinción, para genes con los mismos loci correspondientes. Un cromosoma homólogo se hereda de la madre del organismo; el otro, del padre. Tras la mitosis, las células hijas tienen el número correcto de genes, que son una mezcla de los genes de los dos progenitores.[6]
En los organismos diploides (2n), el genoma está compuesto por un juego de cada par de cromosomas homólogos, en comparación con los organismos tetraploides, que pueden tener dos juegos de cada par de cromosomas homólogos. Los alelos de los cromosomas homólogos pueden ser diferentes, lo que da lugar a fenotipos distintos de los mismos genes. Esta mezcla de rasgos maternos y paternos se ve favorecida por en entrecruzamiento cromosómico que puede ocurrir durante la meiosis, en el que las longitudes de los brazos cromosómicos y el ADN que contienen dentro de un par de cromosomas homólogos se intercambian entre sí.[7]
Historia
A principios del siglo XX, William Bateson y Reginald Punnett estudiaban la herencia genética y observaron que algunas combinaciones de alelos aparecían con más frecuencia que otras. Esos datos e información fueron explorados más a fondo por Thomas Morgan. Utilizando experimentos de cruces de prueba, reveló que, para un solo progenitor, los alelos de los genes cercanos entre sí a lo largo de la longitud del cromosoma se mueven juntos. Utilizando esta lógica, llegó a la conclusión de que los dos genes que estaba estudiando se encontraban en cromosomas homólogos.[4] Más tarde, en la década de 1930, Harriet Creighton y Barbara McClintock estudiaron la meiosis en células de maíz y examinaron los loci genéticos en los cromosomas del maíz. Creighton y McClintock descubrieron que las nuevas combinaciones alélicas presentes en la descendencia y el evento de entrecruzamiento cromosómico estaban directamente relacionados.[4] Esto demostró la recombinación genética intercromosómica.[4]
Estructura
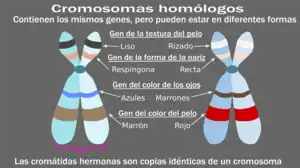
Los cromosomas homólogos son cromosomas que contienen los mismos genes en el mismo orden a lo largo de sus brazos cromosómicos. Hay dos propiedades principales de los cromosomas homólogos:
1) la longitud de los brazos cromosómicos y
2) la ubicación del centrómero.[8]
La longitud real del brazo, de acuerdo con la localización de los genes, es de vital importancia para una alineación adecuada. La colocación del centrómero en el cromosoma puede caracterizarse por cuatro disposiciones principales: metacéntrica, submetacéntrica, acrocéntrica o telocéntrica. Ambas propiedades (es decir, la longitud de los brazos cromosómicos y la colocación del centrómero cromosómico) son los principales factores para crear homología estructural entre cromosomas. Por lo tanto, cuando existen dos cromosomas que contienen relativamente la misma estructura (por ejemplo, el cromosoma 15 materno y el cromosoma 15 paterno), son capaces de emparejarse mediante el proceso de entrecruzamiento cromosómico para formar cromosomas homólogos.[9]
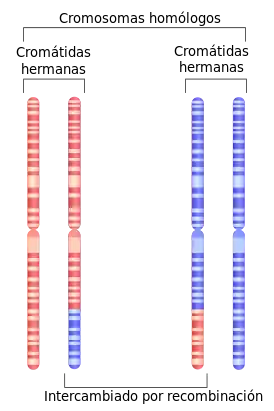
Dado que los cromosomas homólogos no son idénticos y no proceden del mismo organismo, son diferentes de las cromátidas hermanas. Las cromátidas hermanas son el resultado de la replicación del ADN y, por lo tanto, son duplicados idénticos, uno al lado del otro.[10]
En los humanos
Los humanos tienen un total de 46 cromosomas, pero sólo hay 22 pares de cromosomas autosómicos homólogos. El par 23 adicional son los cromosomas sexuales, X e Y.[11]
Hay que tener en cuenta que el par de cromosomas sexuales puede o no ser homólogo, dependiendo del sexo del individuo. Por ejemplo, las hembras contienen XX, por lo que tienen un par homólogo de cromosomas sexuales. Esto significa que las hembras tienen 23 pares de cromosomas homólogos en total (es decir, 22 pares de cromosomas no sexuales (autosomas), 1 par de cromosomas sexuales). Por el contrario, los machos contienen XY, lo que significa que tienen un par de cromosomas sexuales no homólogos como 23º par de cromosomas.
En los humanos, los 22 pares de cromosomas autosómicos homólogos contienen los mismos genes, pero codifican rasgos diferentes en sus formas alélicas, ya que uno se heredó de la madre y otro del padre.[12]
Así pues, los humanos tienen dos juegos de 23 cromosomas en cada célula que contiene un núcleo. Un juego de 23 cromosomas (n) procede de la madre (22 autosomas, 1 cromosoma sexual (sólo X)) y otro juego de 23 cromosomas (n) procede del padre (22 autosomas, 1 cromosoma sexual (X o Y)). En definitiva, esto significa que los humanos son organismos diploides (2n).[4]
Funciones
Los cromosomas homólogos son importantes en los procesos de meiosis y mitosis. Permiten la recombinación y segregación aleatoria del material genético de la madre y el padre en nuevas células.[13]
En la meiosis

La meiosis es una ronda de dos divisiones celulares que da lugar a cuatro células hijas haploides que contienen cada una la mitad del número de cromosomas que la célula madre.[14] Reduce a la mitad el número de cromosomas de una célula germinal separando primero los cromosomas homólogos en la meiosis I y después las cromátidas hermanas en la meiosis II. El proceso de la meiosis I suele ser más largo que el de la meiosis II porque se necesita más tiempo para que la cromatina se replique y para que los cromosomas homólogos se orienten y segreguen correctamente mediante los procesos de apareamiento y sinapsis en la meiosis I.[10] Durante la meiosis, la recombinación genética (por segregación aleatoria) y el entrecruzamiento producen células hijas que contienen cada una combinaciones diferentes de genes codificados por la madre y el padre.[14] Esta recombinación de genes permite la introducción de nuevos emparejamientos de alelos y la variación genética.[4] La variación genética entre los organismos contribuye a la estabilidad de una población al proporcionar una gama más amplia de rasgos genéticos sobre los que puede actuar la selección natural.[4]
Profase I
En la profase I de la meiosis I, cada cromosoma se alinea con su pareja homóloga y se empareja completamente. En la profase I, el ADN ya se ha replicado, por lo que cada cromosoma está formado por dos cromátidas idénticas unidas por un centrómero común.[14] Durante el estadio cigoteno de la profase I, los cromosomas homólogos se emparejan entre sí.[14] Este emparejamiento se produce mediante un proceso de sinapsis en el que el complejo sinaptonemal -un andamiaje proteico- se ensambla y une los cromosomas homólogos a lo largo de sus longitudes.[10] La cohesina se entrecruza entre los cromosomas homólogos y les ayuda a resistir la tracción hasta la anafase.[12] El entrecruzamiento genético, un tipo de recombinación, se produce durante la fase paquiténica de la profase I.[14] Además, con frecuencia se produce otro tipo de recombinación denominada recocido de la cadena dependiente de síntesis (SDSA). La recombinación SDSA implica un intercambio de información entre cromátidas homólogas emparejadas, pero no un intercambio físico. La recombinación SDSA no provoca entrecruzamiento.
En el proceso de entrecruzamiento, los genes se intercambian mediante la ruptura y unión de porciones homólogas de las longitudes de los cromosomas. Las estructuras denominadas quiasmas son el lugar del intercambio.[10] Los quiasmas unen físicamente los cromosomas homólogos una vez que se produce el entrecruzamiento y durante todo el proceso de segregación cromosómica durante la meiosis.[10] Ambos tipos de recombinación funcionan como procesos de reparación de daños en el ADN, en particular las roturas de doble cadena. En el estadio diploteno de la profase I, el complejo sinaptonemal se desensambla antes de permitir la separación de los cromosomas homólogos, mientras que las cromátidas hermanas permanecen asociadas por sus centrómeros.[10]
Metafase I
En la metafase I de la meiosis I, los pares de cromosomas homólogos, también conocidos como bivalentes o tétradas, se alinean en un orden aleatorio a lo largo de la placa metafásica.[14] Esta orientación aleatoria es otra forma que tienen las células de introducir variaciones genéticas. Los husos meióticos que emanan de polos opuestos del huso se unen a cada uno de los homólogos (cada par de cromátidas hermanas) en el cinetocoro.[12]
Anafase I
En la anafase I de la meiosis I, los cromosomas homólogos se separan entre sí. La enzima separasa escinde los cromosomas homólogos para liberar la cohesina que mantenía unidos los brazos cromosómicos homólogos.[12] Esto permite que se liberen los quiasmas y que los homólogos se desplacen a polos opuestos de la célula.[12] Los cromosomas homólogos se segregan al azar en dos células hijas que se someterán a la meiosis II para producir cuatro células germinales hijas haploides.[4]
Meiosis II
Tras la separación de las tétradas de cromosomas homólogos en la meiosis I, se separan las cromátidas hermanas de cada par. Las dos células hijas haploides (el número de cromosomas se ha reducido a la mitad: antes había dos juegos de cromosomas, pero ahora cada juego existe en dos células hijas diferentes que han surgido de la única célula madre diploide por meiosis I) resultantes de la meiosis I sufren otra división celular en la meiosis II, pero sin otra ronda de replicación cromosómica. Las cromátidas hermanas de las dos células hijas son separadas durante la anafase II por las fibras del huso nuclear, dando lugar a cuatro células hijas haploides.[4]
En la mitosis
Los cromosomas homólogos no funcionan igual en la mitosis que en la meiosis. Antes de cada división mitótica que sufre una célula, los cromosomas de la célula madre se replican. Normalmente, los cromosomas homólogos de la célula no se emparejan ni se recombinan genéticamente.[14] En su lugar, las cromátidas hermanas se alinearán a lo largo de la placa metafásica y se separarán de la misma forma que en la meiosis II, al ser separadas en sus centrómeros por los husos mitóticos nucleares.[15] Si se produce algún cruzamiento entre cromátidas hermanas durante la mitosis, no se produce ningún nuevo genotipo recombinante.[4]
En células somáticas
En la mayoría de los contextos, el emparejamiento homólogo se refiere a las células de la línea germinal, pero también tiene lugar en las células somáticas. Por ejemplo, en los humanos, las células somáticas tienen un emparejamiento homólogo muy estrechamente regulado (separado en territorios cromosómicos y emparejamiento en loci específicos bajo el control de la señalización del desarrollo). Sin embargo, en otras especies (sobre todo en Drosophila) el emparejamiento homólogo es mucho más frecuente. En Drosophila, el emparejamiento homólogo favorece un fenómeno de regulación génica denominado transvección, en el que un alelo de un cromosoma afecta a la expresión del alelo homólogo del cromosoma homólogo.[16] Una función notable de este fenómeno es la regulación sexualmente dimórfica de los genes ligados al cromosoma X.[17]
Problemas
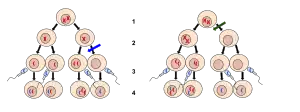
2. Meiosis II
3. Fecundación
4. Cigoto.
La no disyunción se produce cuando los cromosomas no se separan con normalidad, lo que da lugar a una ganancia o pérdida de cromosomas. En la imagen de la izquierda, la flecha azul indica que la no disyunción tiene lugar durante la meiosis II. En la imagen de la derecha, la flecha verde indica la no disyunción durante la meiosis I.
La segregación incorrecta de los cromosomas tiene graves consecuencias. Una segregación defectuosa puede provocar problemas de fertilidad, muerte embrionaria, defectos congénitos y cáncer.[18] Aunque los mecanismos de emparejamiento y adhesión de cromosomas homólogos varían de un organismo a otro, su correcto funcionamiento es imprescindible para que el material genético final se clasifique correctamente.[18]
No disyunción
La correcta separación de los cromosomas homólogos en la meiosis I es crucial para la separación de las cromátidas hermanas en la meiosis II.[18] Un fallo en la separación se conoce como no disyunción. Existen dos tipos principales de no disyunción: la trisomía y la monosomía. La trisomía se produce por la presencia de un cromosoma adicional en el cigoto en comparación con el número normal, y la monosomía se caracteriza por la presencia de un cromosoma menos en el cigoto en comparación con el número normal. Si esta división desigual se produce en la meiosis I, ninguna de las células hijas tendrá una distribución cromosómica adecuada y pueden producirse efectos atípicos, como el síndrome de Down.[19] La división desigual también puede producirse durante la segunda división meiótica. La no disyunción que se produce en esta fase puede dar lugar a células hijas normales y a células deformes.[8]
Otros usos
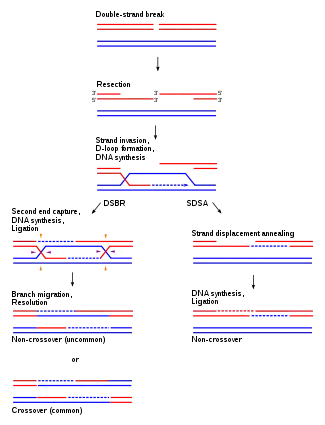
Aunque la función principal de los cromosomas homólogos es su uso en la división nuclear, también se utilizan en la reparación de roturas de doble cadena del ADN.[20] Estas roturas de doble cadena pueden producirse en el ADN en replicación y suelen ser el resultado de la interacción del ADN con moléculas dañinas naturales, como las especies reactivas del oxígeno. Los cromosomas homólogos pueden reparar estos daños alineándose con cromosomas de la misma secuencia genética.[20] Una vez que los pares de bases se han emparejado y orientado correctamente entre las dos cadenas, los cromosomas homólogos llevan a cabo un proceso que es muy similar a la recombinación o entrecruzamiento que se observa en la meiosis. Parte de la secuencia de ADN intacta se solapa con la del cromosoma dañado. Las proteínas y los complejos de replicación se dirigen entonces al lugar del daño, lo que permite la reparación y la replicación adecuada. Gracias a este funcionamiento, las roturas de doble cadena pueden repararse y el ADN puede funcionar con normalidad.[20]
Investigación relevante
La investigación actual y futura sobre el tema de los cromosomas homólogos se centra en gran medida en las funciones de diversas proteínas durante la recombinación o durante la reparación del ADN. En un artículo publicado recientemente por Pezza et al. la proteína conocida como HOP2 es responsable tanto de la sinapsis de cromosomas homólogos como de la reparación de roturas de doble cadena mediante recombinación homóloga. La deleción de HOP2 en ratones tiene grandes repercusiones en la meiosis.[21] Otros estudios actuales se centran también en proteínas específicas implicadas en la recombinación homóloga.
Se está investigando la capacidad de los cromosomas homólogos para reparar las roturas de doble cadena del ADN. Los investigadores están estudiando la posibilidad de explotar esta capacidad para la medicina regenerativa.[22] Esta medicina podría ser muy prevalente en relación con el cáncer, ya que se cree que los daños en el ADN contribuyen a la carcinogénesis. Manipular la función reparadora de cromosomas homólogos podría permitir mejorar el sistema de respuesta al daño de una célula. Aunque la investigación aún no ha confirmado la eficacia de este tratamiento, podría convertirse en una terapia útil contra el cáncer.[23]
Véase también
Referencias
- «Cromosoma homólogos. Diccionario médico. Clínica Universidad de Navarra.». www.cun.es. Consultado el 13 de marzo de 2023.
- «Homologous chromosomes». 2. Philadelphia: Saunders/Elsevier. 2008. pp. 815, 821-822. ISBN 1-4160-2255-4.
- «Fundación Instituto Roche - Glosario de genética - Cromosomas homólogos». www.institutoroche.es. Consultado el 13 de marzo de 2023.
- Griffiths JF, Gelbart WM, Lewontin RC, Wessler SR, Suzuki DT, Miller JH (2005). Introduction to Genetic Analysis. Nueva York: W.H. Freeman and Co. pp. 34-40, 473-476, 626-629. ISBN 0-7167-4939-4.
- Fuertes, María de los Ángeles Gama (2004). Biologia i. Pearson Educación. p. 132. ISBN 978-970-26-0509-6. Consultado el 13 de marzo de 2023.
- Fuertes, María de los Ángeles Gama (2004). Biologia i. Pearson Educación. p. 136. ISBN 978-970-26-0509-6. Consultado el 13 de marzo de 2023.
- Campbell NA, Reece JB (2002). Biology. San Francisco: Benjamin Cummings. ISBN 0-8053-6624-5.
- Klug, William S. (2012). Concepts of Genetics. Boston: Pearson. pp. 21-22.
- Klug, William; Michael Cummings; Charlotte Spencer; Michael Pallodino (2009). «Chromosome Mutations: Variation in chromosome number and arrangement». En Beth Wilbur, ed. Concepts of Genetics (9 edición). San Francisco, CA: Pearson Benjamin Cumming. pp. 213-214. ISBN 9780321540980.
- Pollard TD, Earnshaw WC, Lippincott-Schwartz J (2008). Cell Biology (2 edición). Philadelphia: Saunders/Elsevier. pp. 815, 821–822. ISBN 978-1-4160-2255-8.
- Townsend, Courtney M. (2005-09). Sabiston Tratado de Cirugia: Libro con acceso a sitio Web. Elsevier España. p. 28. ISBN 978-84-8174-848-2. Consultado el 13 de marzo de 2023.
- Lodish HF (2013). Molecular cell biolog. New York: W.H. Freeman and Co. pp. 355, 891. ISBN 978-1-4292-3413-9.
- Gregory MJ. «The Biology Web». Clinton Community College – State University of New York. Archivado desde el original el 16 de noviembre de 2001.
- Gilbert SF (2014). Developmental Biology. Sunderland, MA: Sinauer Associates, Inc. pp. 606–610. ISBN 978-0-87893-978-7.
- «The Cell Cycle & Mitosis Tutorial». The Biology Project. University of Arizona. Oct 2004.
- Lewis, E. B. (July 1954). «The Theory and Application of a New Method of Detecting Chromosomal Rearrangements in Drosophila melanogaster». The American Naturalist (en inglés) 88 (841): 225-239. ISSN 0003-0147. S2CID 222327165. doi:10.1086/281833.
- Galouzis, Charalampos Chrysovalantis; Prud’homme, Benjamin (22 de enero de 2021). «Transvection regulates the sex-biased expression of a fly X-linked gene». Science (en inglés) 371 (6527): 396-400. Bibcode:2021Sci...371..396G. ISSN 0036-8075. PMID 33479152. S2CID 231666458. doi:10.1126/science.abc2745.
- Gerton JL, Hawley RS (June 2005). «Homologous chromosome interactions in meiosis: diversity amidst conservation». Nat. Rev. Genet. 6 (6): 477-87. PMID 15931171. S2CID 31929047. doi:10.1038/nrg1614.
- Tissot, Robert; Kaufman, Elliot. «Chromosomal Inheritance». Human Genetics. University of Illinois at Chicago. Archivado desde el original el 10 de octubre de 1999.
- Sargent RG, Brenneman MA, Wilson JH (January 1997). «Repair of site-specific double-strand breaks in a mammalian chromosome by homologous and illegitimate recombination». Mol. Cell. Biol. 17 (1): 267-77. PMC 231751. PMID 8972207. doi:10.1128/MCB.17.1.267.
- Petukhova GV, Romanienko PJ, Camerini-Otero RD (Dec 2003). «The Hop2 protein has a direct role in promoting interhomolog interactions during mouse meiosis». Dev Cell 5 (6): 927-36. PMID 14667414. doi:10.1016/s1534-5807(03)00369-1.
- González F, Georgieva D, Vanoli F, Shi ZD, Stadtfeld M, Ludwig T, Jasin M, Huangfu D (2013). «Homologous Recombination DNA Repair Genes Play a Critical Role in Reprogramming to a Pluripotent State». Cell Reports 3 (3): 651-660. PMC 4315363. PMID 23478019. doi:10.1016/j.celrep.2013.02.005.
- Khanna KK, Jackson SP (2001). «DNA double-strand breaks: Signaling, repair and the cancer connection». Nature Genetics 27 (3): 247-254. PMID 11242102. S2CID 3012823. doi:10.1038/85798.
Otras lecturas
- Gilbert SF (2003). Developmental biolog. Sunderland, Mass.: Sinauer Associates. ISBN 0-87893-258-5.
- OpenStaxCollege (25 Apr 2013). «Meiosis». Rice University.
- Esta obra contiene una traducción derivada de «Homologous chromosome» de Wikipedia en inglés, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
| Control de autoridades |
|
|---|
 Datos: Q849622
Datos: Q849622 Multimedia: Homologous chromosomes / Q849622
Multimedia: Homologous chromosomes / Q849622