Notocorda
La notocorda, notocordio o cuerda dorsal es una estructura embrionaria común en todos los miembros del phylum Chordata. En los vertebrados superiores la notocorda existe transitoriamente y tiene al menos dos funciones importantes. Primero, la notocorda está posicionada centralmente en el embrión con respecto al eje dorso-ventral y el eje izquierda-derecha. Aquí, produce factores secretados que señalizan los tejidos circundantes, proveyendo información de posición y destino.[1] Segundo, la notocorda juega un papel estructural importante. Como tejido, se encuentra más cercano al cartílago y es probable que represente una forma primitiva de cartílago. De acuerdo a esto, la notocorda sirve como esqueleto axial del embrión hasta que otros elementos como las vértebras se formen. Las células de la notocorda producen una vaina gruesa basal de membrana, y retienen materiales hidratados en grandes vacuolas. Estas vacuolas le permiten a la célula ejercer presión en contra de las paredes de la vaina, lo que le confiere a la notocorda sus propiedades estructurales.[1] En algunos clados de vertebrados, como los agnatos y peces primitivos, la notocorda persiste toda la vida. En vertebrados superiores, la notocorda contribuye a la formación del núcleo pulposo de los discos intervertebrales.[1]
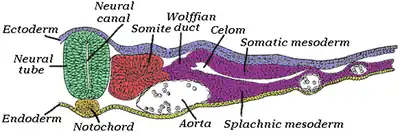
Formación de la notocorda
Urocordados
En los urocordados la notocorda es formada a partir de ciertas células dorsales medias de la pared del arquenterón. Estas se disponen para formar un cordón elongado de células que luego pasan a encontrarse entre la pared del arquenterón y la ranura medular.[2] En este grupo, la notocorda solo se presenta en la cola durante su estadio larval; posteriormente se fijan al sustrato y pierden la cola.
Cefalocordados
La notocorda es la principal estructura de soporte o esquelética de los cefalocordados o anfioxos, la cual se encuentra adelgazada en sus extremos y se extiende a lo largo de toda la longitud del cuerpo en el plano medio. Se encuentra inmediatamente por encima del tubo digestivo y entre los miómeros izquierdos y derechos. Está formada por un tejido especial celular llamado tejido notocordal, el cual está compuesto por células grandes vacuolizadas repletas de líquido, las cuales le dan turgencia y resistencia a la notocorda.[3] Esta estructura en los cefalocordados es formada al tiempo que se forma el tubo neural. Las células cordales que ocupan la parte dorsal media del arquenterón sufren estiramiento, enrollamiento y separaciones para formar la notocorda. La separación del material notocordal empieza en el extremo anterior del embrión como una evaginación dorsal del endodermo y continua lentamente en dirección antero-posterior. Finalmente, se convierte en un cilindro sólido de células vacuoladas que contienen una sustancia gelatinosa de soporte. Posteriormente, la notocorda es rodeada por tejido conectivo fibroso llamado la vaina notocordal.[2] En el anfioxo, único caso en los cordados, la notocorda se extiende anteriormente, por delante de la vesícula cerebral anterior.[3]
Vertebrados
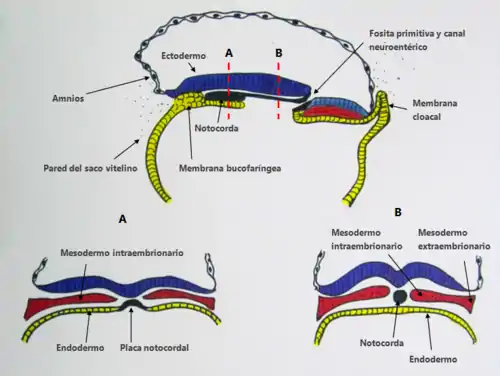
En el embrión, existen dos zonas en las que no existe mesodermo, denominadas membrana bucofaríngea (cefálicamente) y membrana cloacal (caudalmente).[4] Las células prenotocordales se invaginan en la región del nódulo primitiva y migran directamente en dirección cefálica hasta alcanzar la placa precordal (ubicada cerca a la membrana bucofaríngea). Estas células prenotocordales se intercalan en el hipoblasto de manera que durante un breve periodo la línea media del embrión está formada por dos capas celulares que constituyen la placa notocordal.[5] A medida que el hipoblasto es reemplazado por células endodérmicas que se desplazan hacia la línea primitiva, las células de la placa notocordal proliferan y se desprenden de endodermo formando un cordón macizo llamado notocorda definitiva, la cual se encuentra por debajo del tubo neural y que se extiende desde la placa precordal en la parte craneal, hacia el esbozo caudal en la parte caudal. La notocorda definitiva sirve de base para el esqueleto axial, establece la línea media y envía señales moleculares esenciales para la inducción del tubo neural, las somitas y otras estructuras adyacentes.[5] La elongación de la notocorda es entonces, un proceso dinámico en donde primero se forma el extremo cefálico y las regiones caudales se agregan a medida que la línea primitiva adopta una posición más caudal, es por esto que algunas partes de la notocorda definitiva se establecen primero en la región cefálica.[5] La formación de la notocorda, las somitas y los somitómeros se da gracias a la dorsalización del mesodermo craneal por acción de la cordina (activada por el factor de transcripción goosecoid), la nogina, y la folistatina, los cuales antagonizan la actividad de BMP4 (Proteína morfogenética ósea) encargada de la ventralización del mesodermo (en conjunto con el TGF-β (Factor de crecimiento transformador β)), lo que conlleva a la dorsalización de este.[5]
Después de que la notocorda ha proporcionado la integridad axial del embrión temprano y ha inducido la formación del tubo neural ventral, la mayor parte de esta degenera por apoptosis. Esta apoptosis probablemente es inducida por fuerzas mecánicas. Luego de que se forman los cuerpos vertebrales, entre estos las células notocordales forman parte de los discos intervertebrales, los núcleos pulposos. Estos son los discos que se “deslizan o hernian” en ciertas lesiones de la espalda.[6]
Funciones en el desarrollo
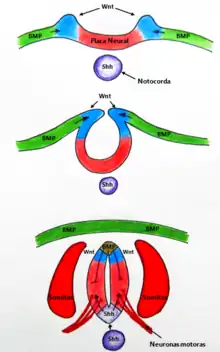
Establecimiento del Patrón Ventral del Tubo Neural
La especificación del eje dorsoventral de tubo neural es inducida por dos factores paracrinos principales. Uno de ellos es la proteína Sonic hedgehog, originada desde la notocorda y posteriormente secretada por las células del tubo neural ventral en diferenciación. Esta proteína induce a las células de bisagra mediales a convertirse en placa de piso o del suelo del tubo neural (floor-plate). Este factor paracrino interacciona con la proteína BMP4 proveniente del ectodermo dorsal. Las células cercanas a la notocorda, reciben altas concentraciones de Sonic Hedgehog gracias a la cual se sintetizan factores de transcripción que generan la conversión a neuronas ventrales.[6] Experimentos durante la primera parte del siglo XX por Holtfreter demostraron la importancia de la notocorda en la polaridad dorsoventral del tubo neural. Aislamientos del tubo neural de tejidos circundantes resultaron en un tubo indiferenciado. Sin embargo, cuando trasplantó una nueva notocorda al lado más dorsal, se indujo una segunda placa de piso y diferenciación a neuronas motoras en el tubo neural dorsal.[7]
Establecimiento del Eje Izquierda-Derecha
La notocorda también ejerce funciones en el establecimiento del eje izquierda-derecha o lateral. El FGF8 secretado por el nódulo primitivo causa la expresión de nodal en el mesodermo del lado izquierdo del embrión. A su vez, nodal inicia una cascada de expresión genética en el lado izquierdo de la placa neural, la cual es bloqueada en el lado derecho por la expresión de sonic hedgehog en la notocorda y de lefty1. Adicionalmente, la notocorda expresa el gen Brachyury, el cual codifica para un factor de transcripción esencial para la expresión de nodal, lefty1 y lefty2.[5]
Determinación del Esclerotoma
La notocorda interviene en la especificación de las somitas. Secretado desde la notocorda y desde la placa del piso del tubo neural, la porción ventromedial del somita es inducida a convertirse en esclerotoma. Posteriormente estas células expresan un nuevo factor de transcripción Pax1, que es requerido para su diferenciación hacia cartílago y cuya presencia es necesaria para la formación de las vértebras.[6] Por otra parte, parece ser que la notocorda induce a las células mesenquimáticas de su alrededor a secretar epimorfina y esta epimorfina atrae a las células del esclerotoma hacia la región alrededor de la notocorda y el tubo neural.[6]
Referencias
- Stemple, D.L (2005) Structure and function of the notochord: an essential organ for chordate development. Development,132, pp 2503-2512
- Sastry, K. & Vineeta, S. (2010) Developmental Biology. Meerut: Rastogi Publicarions
- Parker, T.J. & Haswell, W.A. (2010) Biología de Cordados (Vol II). Barcelona: Editorial Reverté
- Henríquez (S.f.) Embriología del Sistema Nervioso Central. Universidad de la Frontera. Extraído el 20 de noviembre de 2012 de: Archivado el 4 de noviembre de 2012 en Wayback Machine.
- Sadler, T.W. (2008) Langman Embriología Médica: Con Orientación Clínica (10a Edición). Buenos Aires: Médica Panamericana
- Gilbert, S. F. (2006) Biología del Desarrollo (7ª Edición). Buenos Aires: Médica Panamericana
- Sanes, D.H., Reh, T.A. & Harris, W.A. (2006) Development of the Nervous System (2nd Edition) London: Elsevier Inc