Depresión a largo plazo
La depresión a largo plazo (DLP)[1] en neurofisiología, es un tipo de plasticidad neuronal en el que se hay una reducción de la eficacia de la sinapsis neuronal. La expresión de DLP depende de la actividad de las neuronas que conforman la sinapsis. La DLP dura horas o más.
La DLP tiene lugar en diversas áreas del sistema nervioso central con diferentes mecanismos dependiendo del área del cerebro y del periodo del desarrollo del mismo. Por ejemplo, la DLP en el hipocampo y el cerebelo son las que más se han estudiado, pero también existen otras áreas del cerebro en las cuales la DLP existe.[2] Se ha comprobado que la DLP ocurre en diferentes tipos de neuronas que liberan varios neurotransmisores, excitatorios o inhibitorios. Puede resultar de estímulos sinápticos fuertes, como ocurre en células de Purkinje del cerebelo, o de estímulos sinápticos débiles pero persistentes como ocurre en el hipocampo. Se estima que la DLP resulta del decremento en la densidad de receptores postsinápticos,[3] aunque también puede ser influenciada por decrecimiento en la liberación de neurotransmisores.[4] Tradicionalmente, la DLP en el cerebelo se ha asociado con el aprendizaje motriz en la corteza, sin embargo, recientes experimentos no apoyan esta hipótesis.[3] En cambio en el hipocampo, la DLP puede ser importante para deshacerse de trazos de memorias viejas.[5][6]
La DLP es uno de los diferentes procesos que sirve para debilitar de manera selectiva sinapsis específicos de manera que se pueda hacer uso constructivo de la fortaleza de la sinapsis causada por la PLP. Esto es necesario porque, si se continúa incrementando en fuerza, la sinapsis llegaría a su máximo nivel de eficacia, que en consecuencia inhibiría procesamiento de información nueva.[7]
La DLP en el cerebelo
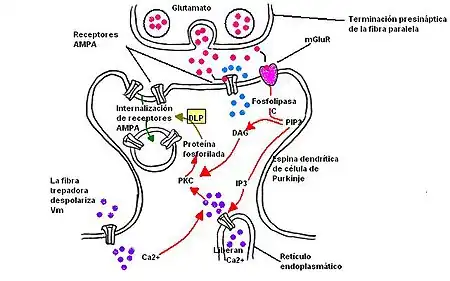
La DLP en el cerebelo ocurre en la sinapsis de las células de Purkinje, ellas reciben dos formas de estimulación excitatoria: una de parte de fibras trepadoras y la otra de fibras paralelas. La DLP disminuye la eficacia de la transmisión de las fibras paralelas.[8] Para que DLP ocurra ambas fibras requieren ser activadas simultáneamente. Por una vía, fibras paralelas sueltan glutamato para activar a los receptores No NMDA, principalmente a los AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) y kainato, y también a los receptores metabotrópicos de glutamato en la parte postsináptica de las célula de Purkinje. Cuando glutamato se adhiere al receptor AMPA, la membrana se despolariza. Pero cuando glutamato se adhiere al receptor metabotrópico, diacilglicerol (DAG) es producido junto con inositol trifosfato (IP3); un segundo mensajero de la transducción de señal celular. En la vía iniciada por la activación de las fibras trepadoras, calcio entra en la célula postsináptica a través de los canales iónicos dependientes de voltaje, incrementando niveles de calcio intracelular. Juntos, DAG y IP3 aumentan la concentración de la concentración de calcio al activar la proteína quinasa C (PKC, del inglés protein kinase C). La PKC fosforiliza receptores AMPA, esto da como resultado la internalización de los receptores de AMPA. Con esta pérdida de receptores AMPA la respuesta de la célula de Purkinje postsináptica es menos sensible a la liberación de glutamato desde las terminaciones presinápticas de las fibras paralelas.[8]
La DLP en el hipocampo
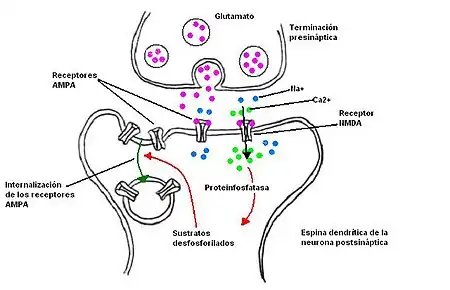
La DLP en estas dos regiones depende del tiempo y de la frecuencia en la corriente de entrada de calcio y se da a cabo cuando pequeños incrementos en el nivel de calcio postsináptico ocurren.[9] Cuando el nivel de entrada de calcio es menor al umbral da como resultado la DLP. El nivel del umbral en la región CA1 fluctúa y depende del historial de la sinapsis. Si la sinapsis ha sido sujetada a PLP, el umbral incrementa, dando así mayor probabilidad de que la entrada de calcio cause DLP. De esta forma el sistema de retroalimentación negativa mantiene una plasticidad sináptica. La activación de receptores NMDA de tipo A es requerida para la entrada de calcio a células postsinápticas de la región CA1.[10] Cambio en el voltaje proporciona un control gradual de calcio postsináptico mediante el control de entrada de calcio dependientes de NMDAR, el cual es responsable en iniciar la DLP. La PLP se debe parcialmente a la activación de la CaMKII, que fosforila las proteínas diana. La DPL es sin embargo el resultado de la activación de fosfatasa dependiente de Ca2+, que separan los grupos fosfato de estas moléculas diana. Los diferentes efectos del Ca2+ durante la DPL y la PLP se deben a la activación selectiva de cinasas y proteinfosfatasas por los altos o bajos niveles del Ca2+. Es posible que las cinasas y las fosfatas fosforilen y desfosforilen el mismo conjunto de proteínas reguladoras para controlar la eficacia de la transmisión en la sinapsis colateral de Schaffer-CA1. La DLP se asocia a la pérdida de receptores AMPA sinápticos porque éstos se internalizan en la célula postsináptica.[11]
Los tipos de DLP
La depresión a largo plazo (DLP) se puede dividir en homosináptica y heterosináptica. La DLP homosináptica se refiere a las sinapsis individuales que se activan por un estímulo de baja frecuencia. En otras palabras, esta forma de DLP depende solamente de la actividad, porque el evento causante del debilitamiento sináptico ocurre en la misma sinapsis que se está activando4. La DLP heterosináptica, en contraste, ocurre en las sinapsis que están inactivas. El debilitamiento de la sinapsis se debe a disparos de interneuronas que están modulando la sinapsis y no a la actividad de las neuronas presináptica y postsináptica. Por ende, esta forma de DLP impacta a sinapsis cercanas a aquellas que están recibiendo los potenciales de acción.[12] Un fenómeno heterosináptico es un evento que ocurre en las uniones (sinapsis) que forma conexiones entre neuronas. En otras palabras es aquel que afecta directamente al elemento presináptico y postsináptico de la sinapsis en cuestión. Este término es usado en relación a depresión a largo plazo y potenciación a largo plazo en neuronas activas. Por ejemplo, depresión a largo plazo heterosináptica ocurre cuando actividad neutral que involucre una de las neuronas sinapsis cause debilitamiento de la sinapsis de otra neurona. Interacciones homosinápticas son eventos que ocurren en la sinapsis donde se encuentran las neuronas. Eventos homosinápticos es aquel que afecta indirectamente al elemento presináptico y postsináptico de la sinapsis en cuestión. Este término es usado en relación con depresión a largo plazo y potenciación a largo plazo en neuronas activas. Por ejemplo, potenciación a largo plazo homosináptico ocurre en sinapsis que conectan dos neuronas disparando al mismo tiempo.
Referencias
- Ito M. The molecular organization of cerebellar long-term depression. Nat Rev Neurosci. 2002 Nov;3(11):896-902. Review. PubMed PMID 12415297
- Massey PV, Bashir ZI (April 2007). "Long-term depression: multiple forms and implications for brain function". Trends Neurosci. 30 (4): 176–84. doi:10.1016/j.tins.2007.02.005. PMID 17335914
- Schonewille M, Gao Z, Boele HJ, Vinueza Veloz MF, Amerika WE, Simek AA, De Jeu MT, Steinberg JP, Takamiya K, Hoebeek FE, Linden DJ, Huganir RL, De Zeeuw CI. Reevaluating the Role of LTD in Cerebellar Motor Learning. Neuron. 2011 Apr 14;70(1):43-50. PubMed PMID 21482355
- Zhao Y, Tzounopoulos T. Physiological activation of cholinergic inputs controls associative synaptic plasticity via modulation of endocannabinoid signaling. J Neurosci. 2011 Mar 2;31(9):3158-68. PubMed PMID 2136802
- Nicholls RE, Alarcon JM, Malleret G, Carroll RC, Grody M, Vronskaya S, Kandel ER (April 2008). "Transgenic mice lacking NMDAR-dependent LTD exhibit deficits in behavioral flexibility". Neuron 58 (1): 104–17. doi:10.1016/j.neuron.2008.01.039. PMID 18400167
- Malleret G, Alarcon JM, Martel G, Takizawa S, Vronskaya S, Yin D, Chen IZ, Kandel ER, Shumyatsky GP (Mar 2010). "Bidirectional regulation of hippocampal long-term synaptic plasticity and its influence on opposing forms of memory". J Neurosci. 30 (10): 3813–25. doi:10.1523/JNEUROSCI.1330-09.2010. PMID 20220016
- Purves D (2008). Neuroscience (4th ed.). Sunderland, Mass: Sinauer. pp. 197–200. ISBN 0-87893-697-1
- Purves D (2008). Neuroscience (4th ed.). Sunderland, Mass: Sinauer. pp. 197–200. ISBN 0-87893-697-1
- Bear MF (July 1995). "Mechanism for a sliding synaptic modification threshold". Neuron 15 (1): 1–4. doi:10.1016/0896-6273(95)90056-X. PMID 7619513. «Copia archivada». Archivado desde el original el 23 de junio de 2010. Consultado el 26 de octubre de 2009.
- Blanke ML, VanDongen AM (2008). "Activation Mechanisms of the NMDA Receptor". In VanDongen AM. Biology of the NMDA Receptor (Frontiers in Neuroscience). Boca Raton: CRC. ISBN 1-4200-4414-1
- Purves, Augustine, Fitzpatrick, Hall, LaMantia, Williams: Neurociencia.pp658,659,660,661.
- Escobar ML, Derrick B (2007). "Long-Term Potentiation and Depression as Putative Mechanisms for Memory Formation". In Bermudez-Rattoni F. Neural plasticity and memory: from genes to brain imaging. Boca Raton: CRC Press. ISBN 0-8493-9070-2