Célula ependimaria
Las células ependimogliales,[1] ependimarias o tanicitos, forman parte del conjunto de células neurogliales del tejido nervioso. Recubren los ventrículos del cerebro y el conducto ependimario y muestran múltiples subtipos celulares identificables por su morfología. Las células ependimarias poseen un potencial latente de células madre neurales.
| Célula ependimaria | ||
|---|---|---|
 Canal central de la médula espinal, células ependimarias | ||
| Nombre y clasificación | ||
| Latín | Ependyma | |
| TA | A14.1.00.022 | |
| Gray | pág.829 | |
| Información anatómica | ||
| Región | epéndimo | |
| Sistema | Sistema nervioso | |
|
| ||
Embriología
Derivan del ectodermo neural del sistema nervioso en desarrollo, por lo tanto no son células epiteliales, porque carecen de membrana basal y de uniones intercelulares características del tejido epitelial.
Son descendientes de las células madre neuroepiteliales embrionarias, localizadas en el tubo neural ventral.
Estructura

Las ependimarias son células de forma cilíndrica a cuboide, que recubren los ventrículos del cerebro y el conducto central de la médula espinal.[2]
La arquitectura microscópica de las células ependimarias ciliadas que contactan con la luz, es heterogénea con células que muestran una morfología típica de células ependimarias cuboideas y otras con una morfología de tanicitos.
Además es visible un tercer tipo de célula ependimaria menos numerosa, la célula ependimaria radial. Las radiales comparten la morfología del citoplasma, y a menudo del núcleo, con las otras células, pero poseen un proceso basal largo. Estas células radiales se encuentran en el polo dorsal o en el ventral de la capa ependimaria, con su proceso basal orientado a lo largo del eje dorso-ventral.[3]
Ultraestructura
Su citoplasma contiene una gran cantidad de mitocondrias.
Presentan haces de filamentos intermedios nestina y vimentina, que se asocian habitualmente con células neurales inmaduras.
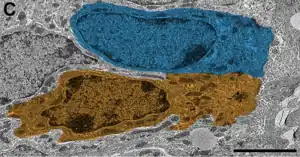
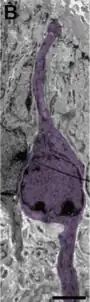
El análisis ultraestructural reveló morfología heterogénea entre estas células ciliadas que contactan con la luz. Las células cuboideas muestran núcleo oval, citoplasma claro y de 1-3 cilias.
Los tanicitos tienen núcleo irregular, citoplasma oscuro y prolongaciones.
Las células radiales presentan núcleo oval e irregular, citoplasma claro, de 1-3 cilias y prolongaciones largas.[3]
En algunas regiones estas células son ciliadas, una característica que facilita el movimiento del líquido cefalorraquídeo.
En el embrión, las prolongaciones que surgen del cuerpo celular llegan a la superficie del encéfalo, pero en la persona adulta las prolongaciones son reducidas y sólo tiene terminaciones cercanas.
En los lugares donde el tejido neural es delgado, esta células forman una membrana limitante interna que recubre el ventrículo cerebral y una membrana limitante externa debajo de la piamadre.
A nivel de los ventrículos cerebrales, estas células sufren modificaciones y forman los plexos coroideos, que tiene como misión secretar y conservar la composición química del líquido cefalorraquídeo.
Tanicitos
Ep= ependimocitos.
III-V= tercer ventrículo.
Los tanicitos son células ependimarias especializadas de contacto entre el tercer ventrículo y la eminencia media del hipotálamo basal.
Su polo proximal se encuentra dentro de la pared del tercer ventrículo y su proceso celular largo se proyecta hacia el hipotálamo ventromedial.
Tipos de tanicitos
Hay cuatro tipos de tanicitos: α1, α2, β1 y β2. Los tanicitos α2 y β1 se localizan en la pared lateral inferior del tercer ventrículo; tienen procesos celulares extendidos que contactan las neuronas en el núcleo arqueado (AN), en particular las neuronas del neuropéptido Y (NPY) y las neuronas de pro-opiomelanocortina (POMC).
Los tanicitos β2 forman la barrera del líquido cefalorraquídeo de la eminencia media (ME) y sus procesos extendidos entran en contacto con vasos desprovistos de la barrera hematoencefálica y a veces están en contacto directo con microvasos presentes en la ME.[4] [5]
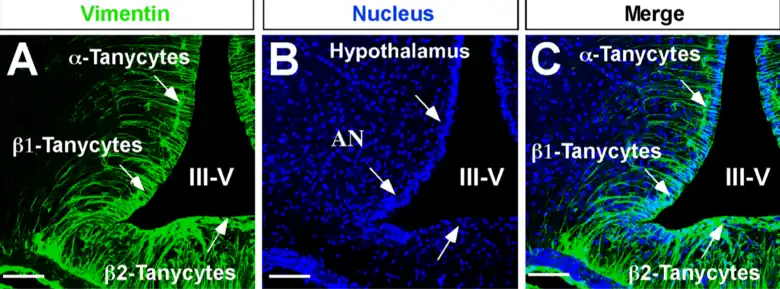
Tienen funciones conocidas, y se les ha atribuido un papel de transporte de sustancias entre ambas estructuras.
Recambio celular de ependimocitos
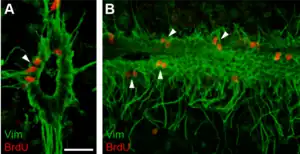
Las células ependimarias (EpC), exhiben una actividad mitótica basal relativamente baja.[6] El uso de BrdU, que se incorpora al ADN en las células en la fase S, dio como resultado el marcado de 16-24% de las células ependimarias en un mes.
En los mamíferos adultos se producen nuevas neuronas de manera continua.[7] La capa de células ependimaria forma nichos de neurogénesis adulta en la zona subventricular (ZSV), en todos los puntos a lo largo del neuroeje.[8]
El daño mecánico del canal central desencadena un fuerte aumento reactivo de la proliferación de las células ependimarias (EpC). Esas EpC reactivas de mamíferos, reexpresan las propiedades de las células madre neurales después de una lesión de la médula espinal.[6]
Referencias
- OMS,OPS (ed.). «Células Ependimogliales». Biblioteca virtual de salud, Descriptores en Ciencias de la Salud. Consultado el 21 de octubre de 2020.
- Gartner L. (2002). «cap9: Tejido nervioso». Texto atlas de histología (2da edición). issuu.com. pp. 190-191. Consultado el 20 de octubre de 2020.
- Meletis K.; Barnabé-Heider F.; Carlén M.; Evergren E.; Tomilin N.; Shupliakov O.; Frisén J. (2008). Spinal Cord Injury Reveals Multilineage Differentiation of Ependymal Cells 6 (7). PLoS Biology . p. e182. doi:10.1371/journal.pbio.0060182. Consultado el 21 de octubre de 2020.

- Elizondo-Vega R., Cortes-Campos C., Barahona M.J., Oyarce K.A., Carril C.A., García-Robles M.A. (2015). «The role of tanycytes in hypothalamic glucosensing». Journal of Cellular and Molecular Medicine 19 (7): 1471-1482. doi:10.1111/jcmm.12590. Consultado el 19 de octubre de 2020.

- Salgado M, Tarifeño-Saldivia E, Ordenes P, Millán C, Yañez MJ, Llanos P, et al (2014). «Dynamic Localization of Glucokinase and Its Regulatory Protein in Hypothalamic Tanycytes». PLoS ONE 9 (4): e94035. doi:10.1371/journal.pone.0094035. Consultado el 19 de octubre de 2020.
- Lacroix S.; Hamilton L.K.; Vaugeois A.; Beaudoin S.; Breault-Dugas C.; Pineau I.; Lévesque S.A.; Grégoire C-A.; Fernandes K.J.L. (2014). «Central Canal Ependymal Cells Proliferate Extensively in Response to Traumatic Spinal Cord Injury but Not Demyelinating Lesions». PLoS ONE 9 (1): e85916. doi:10.1371/journal.pone.0085916. Consultado el 22 de octubre de 2020.
- Samuel Weiss; Christine Dunne; Jennifer Hewson; Cheryl Wohl; Matt Wheatley; Alan C. Peterson; Brent A. Reynolds (1996). «Multipotent CNS Stem Cells Are Present in the Adult Mammalian Spinal Cord and Ventricular Neuroaxis». Journal of Neuroscience 16 (23): 7599-7609. doi:10.1523/JNEUROSCI.16-23-07599.1996.
- Gouazé A.; Brenachot X.; Rigault C.; Krezymon A.; Rauch C.; Nédélec E.; Lemoine A., Gascuel J., Bauer S., Pénicaud L., Benani A. (2013). «Cerebral Cell Renewal in Adult Mice Controls the Onset of Obesity». PLoS ONE 8 (8): e72029. doi:10.1371/journal.pone.0072029. Consultado el 20 de octubre de 2020.
 Datos: Q28646122
Datos: Q28646122