Espina dendrítica
Una espina dendrítica, o espina, es una pequeña protuberancia en la membrana del árbol dendrítico de ciertas neuronas donde, típicamente, se produce la sinapsis con un botón axonal de otra neurona, y en ocasiones contactan varios axones. Aunque su morfología es muy variada, una gran parte de las espinas tiene forma de bulbo, la cabeza de la espina, y éste está conectado por un cuello, más fino, al tronco de la dendrita. Esta geometría permite aislar ligeramente las señales eléctricas y químicas de las sinapsis, y a su vez sus propiedades, de las colindantes. Las dendritas de una neurona pueden contener de cientos a miles de espinas. Además de incrementar la superficie de las dendritas y permitir un mayor número de contactos entre neuronas, se considera que las espinas forman la base en la que se sustenta el almacenamiento de la memoria.
.jpg.webp)
Historia
Su descubrimiento y nombre se debe a Santiago Ramón y Cajal durante el estudio de células de Purkinje en el cerebelo de palomas, publicado en Estructura de los centros nerviosos de las aves.[1] Aunque las espinas habían sido observadas anteriormente, se creía que eran artefactos del método de preparación, método de Golgi, por lo que el descubrimiento fue recibido con escepticismo. Ramón y Cajal propuso entonces que las espinas podían servir como los lugares de contacto entre neuronas. La prueba de esto mismo sólo llegó una vez desarrollada la microscopía electrónica.[2]
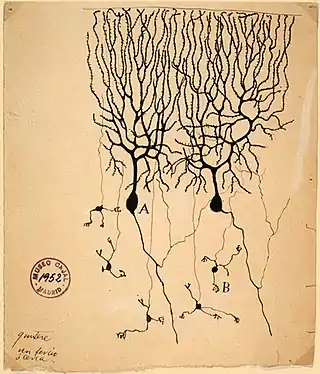
Hasta el desarrollo de la microscopía confocal en tejido vivo, se consideraba que las espinas se formaban durante el desarrollo embrionario y que permanecían estables después del nacimiento. Sin embargo, se pudo demostrar que, de hecho, tienen una estructura movible y dinámica y que se están continuamente renovando, incluso después del nacimiento.[3][4][5]
Características
Estructura
El tamaño de las espinas dendríticas depende de diversos factores, especie, tipo de neurona y su maduración. El volumen de la cabeza puede variar de los 0.01 µm³ a los 0.8 µm³. Ésta suele estar unida a la dendrita mediante un cuello cuyo tamaño es también variable, con longitudes de hasta 2 µm y diámetros del orden de 150 nm. Según su forma se suelen subdividir en diferentes tipos: espina "primordial" o filopodio, espina "delgada", espina "gorda", espina "en hongo", espina "doble" y espina "ramificada". Estudios de microscopía electrónica han demostrado que esta clasificación en realidad es relativamente arbitraria y que hay un continuo de formas. Se ha demostrado así mismo que existe una correlación entre el tamaño de la cabeza de la espina y la fuerza de la sinapsis asociada.
Distribución
Las espinas dendríticas contienen normalmente una sinapsis excitatoria, en contacto con un axón, aunque en ocasiones pueden tener a su vez una o varias sinapsis inhibitorias junto con la excitatoria, contactando axones provenientes de diferentes neuronas. Se suelen encontrar en las dendritas de la mayoría de neuronas principales del cerebro, incluyendo las células piramidales o las células de Purkinje del cerebelo. La densidad de espinas dendríticas depende del tipo de neurona y de la distancia al soma de la célula, siendo del orden de 5 espinas/µm.
Plasticidad
Las espinas dendríticas son plasticas,[6] es decir, pueden cambiar significativamente de forma, volumen e incluso desaparecer en periodos cortos de tiempo. Su mantenimiento y plasticidad depende de diversos factores, entre ellos la actividad de las sinapsis que contienen.[7] Del orden del 10-20% de las espinas de células piramidales del neocortex pueden desaparecer o aparecer espontáneamente en tiempos de segundos o minutos. Así, las espinas más grandes, tipo en hongo, suelen ser las más estables.
Referencias
- Ramón y Cajal, S. Estructura de los centros nerviosos de las aves. Rev. Trim. Histol. Norm. Pat. 1, 1-10 (1888).
- Gray, E. G. (1959). «Electron microscopy of synaptic contacts on dendrite spines of the cerebral cortex». Nature 183: 1592-1593. PMID 13666826. doi:10.1038/1831592a0.
- Dailey, M. E.; Smith, S. J. (1996). «The dynamics of dendritic structure in developing hippocampal slices». J Neurosci 16: 2983-2994.
- Bonhoeffer, T.; Yuste, R. (2002). «Spine motility. Phenomenology, mechanisms, and function». Neuron 35: 1019-1027. PMID 12354393. doi:10.1016/s0896-6273(02)00906-6.
- Yoshihara, Y., De Roo, M. & Muller, D. "Dendritic spine formation and stabilization. Curr Opin Neurobiol (2009).
- Alvarez, V.; Sabatini, B. (2007). «Anatomical and Physiological Plasticity of Dendritic Spines». Annual Review of Neuroscience 30: 79-97. PMID 17280523. doi:10.1146/annurev.neuro.30.051606.094222.
- De Roo, M.; Klauser, P.; Mendez, P.; Poglia, L.; Muller, D. (2007). «Activity-Dependent PSD Formation and Stabilization of Newly Formed Spines in Hippocampal Slice Cultures». Cerebral Cortex 18 (1): 151-161. ISSN 1047-3211. doi:10.1093/cercor/bhm041.
Bibliografía
| Control de autoridades |
|
|---|
 Datos: Q902079
Datos: Q902079 Multimedia: Dendritic spines / Q902079
Multimedia: Dendritic spines / Q902079