Exonucleasa
Las exonucleasas son enzimas que funcionan escindiendo nucleótidos uno a uno a partir del extremo terminal (exo) de una cadena polinucleotídica. Estas enzimas catalizan una reacción de hidrólisis que rompe los enlaces fosfodiester ya sea en el extremo 3' o 5'. Se encuentran estrechamente relacionadas con las endonucleasas, las cuales rompen los enlaces fosfodiester en el medio (endo) de la cadena polinucleotídica. Existen tres tipos de exonucleasas involucradas en la maduración normal del ARNm: la 5' a 3' exonucleasa, la 3' a 5' exonucleasa, que es una proteína independiente y la exonucleasa 3' a 5' específica de poli A.
Tanto en arqueas como en eucariotas, una de las principales rutas de degradación del ARN es llevada a cabo por el complejo exosoma un complejo multiproteico que consiste a grandes rasgos en un gran número de exorribonucleasas 3' a 5'.
Importancia en relación con las polimerasas
La polimerasa de ARN tipo II es conocida por desempeñar una función durante la terminación transcripcional, esta enzima funciona en conjunto con una exonucleasa 5' (en humanos codificada por el gen Xrn2) degradando el transcripto recién formado aguas abajo, dejando el sitio de poliadenilación y disparando simultáneamente la polimerasa. Este proceso implica que la exonucleasa debe ser capaz de atrapar a la polimerasa II y terminar con la transcripción.[1]
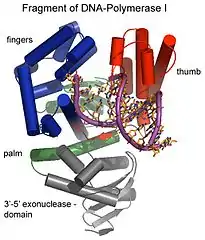
Luego, la Pol I coloca nucleótidos de ADN en el lugar ocupado por el iniciador de ARN que acaba de ser removido. La polimerasa de ADN I también posee actividad exonucleasa 3' a 5', la cual es utilizada en la edición y en el mecanismo de corrección de errores del ADN.
Tipos de exonucleasas presentes en E. coli
En 1971, Lehman descubrió la polimerasa I en E. coli. Desde aquellos días, ha habido muchos nuevos descubrimientos entre los que se incluyen la exonucleasa II, III, IV, V, VII y VIII. Cada tipo de exonucleasa tiene una función o requerimiento específicos.[2]
- La exonucleasa I rompe las hebras de ADN monocatenaro en dirección 3'→5', liberando deoxiribonucleósidos 5'-monofosfato, uno detrás de otro. Sin embargo es incapaz de actuar sobre hebras de ADN sin los grupos terminales 3'-OH ya sea que se encuentren bloqueados por un grupo fosforíl o acetil.[3]
- La exonucleasa II se encuentra asociada a la polimerasa de ADN tipo I, la cual posee actividad exonucleasa 5' que es capaz de cortar el iniciador de ARN ubicado inmediatamente corriente abajo de un sitio de síntesis 5' → 3' del ADN.
- La exonucleasa III posee cuatro actividades catalíticas:
- Actividad exodeoxiribonucleasa 3’ a 5’, la cual es específica para ADN doble cadena
- Actividad RNAsa
- Actividad 3’ fosfatasa
- Actividad endonucleasa de tipo AP (más tarde llamada actividad endonucleasa de tipo II).[4]
- La exonucleasa IV es una hidrolasa que añade una molécula de agua, de modo que es capaz de romper el enlace fosfodiéster de un oligonucleótido para formar un nucleósido 5'-monofosfato. Esta exonucleasa requiere de Mg2+ para su funcionamiento y trabaja a temperaturas mayores que la exonucleasa I[5]
- La exonucleasa V es una enzima hidrolítica con actividad exonucleasa 3’ a 5’ que cataliza la degradación tanto de ADN doble cadena como de ADN de cadena simple, requiere de Ca2+ para su funcionamiento y es extremadamente importante en el proceso de recombinación homóloga.[6]
- La exonucleasa VIII es una proteína dimérica con actividad 5’ a 3’ que no requiere de ATP, ni brechas ni saltos en la cadena para actuar, aunque si requiere de un grupo 5'OH libre para desempeñar su función.
Descubrimientos en humanos
Las endonucleasas de tipo 3' a 5' son conocidas por desempeñar un papel fundamental para el correcto procesamiento del transcripto primario de ARNm para histonas, en el cual las riboproteína U7 perteneciente a las riboproteínas nucleares pequeñas (snRNP) dirige el proceso de ruptura simple. A continuación elimina el siguiente producto de escisión, la exonucleasa 5' a 3' continúa quitando nucleótidos al ARN hasta que es completamente degradado.[7] Esto permite la reutilización de los nucleótidos. La exonucleasa 5'a 3' está relacionada con la actividad de escisión postranscripcional que actúa como precursora para el desarrollo de extremos 5' desprotegidos, de modo que la exonucleasa quede capacitada para degradar el producto de escisión aguas abajo. Esto da inicio a la terminación transcripcional ya que no es conveniente que las cadenas de ADN o ARN se acumulen en el organismo.[8]
Descubrimientos en levaduras
El CCR4-NOT es un complejo regulatorio general presente en levaduras. Se ha encontrado que este complejo se encuentra relacionado con el metabolismo del ARNm, tanto con la iniciación de la transcripción, como con la degradación del mismo. Se ha demostrado también que el complejo CCR4 posee actividad exonucleasa 3' a 5' tanto para ARN como para ADN monocatenario.[9] Otro componente emparentado con el complejo CCR4 es la proteína CAF1, la cual ha demostrado tener actividad 3' a 5' o 5' a 3' en Mus musculus y en Caenorhabditis elegans.[10] Esta proteína no ha sido encontrada en levaduras, lo que sugiere que es probable que CCR4 posea un dominio exonucleasa anormal como la que se observa en los metazoos.[11] Las levaduras contienen además las exonucleasas Rat1 y Xrn1. La Rat1 funciona de la misma manera que la Xrn2 humana, mientras que la actividad de la Xrn1 se encuentra en el citoplasma donde degrada ARN ribosomal de tipo pre-5.8s y 25s en ausencia de Rat1.[12][13]
Enlaces externos
- MeSH: Exonucleases (en inglés)
Referencias
- Hage A EL, et al. (2008). «Efficient termination of transcription by RNA polymerase I requires the 5′ exonuclease Rat1 in yeast». Genes Dev. 22 (8): 1068-081. PMC 2335327. PMID 18413717. doi:10.1101/gad.463708.
- Paul D. Boyer (1952). The Enzymes (1st edición). Academic Press. p. 211. ISBN 0-12-122723-5.
- Lehman IR, Nussbaum AL (agosto de 1964). «The deoxyribonucleases of Escherichia Coli. V. on the specificity of exonuclease I (Phosphodiesterase)». J. Biol. Chem. 239 (8): 2628-36. PMID 14235546. Archivado desde el original el 29 de mayo de 2020. Consultado el 5 de septiembre de 2012.
- Rogers SG, Weiss B (1980). «Exonuclease III of Escherichia coli K-12, an AP endonuclease». Meth. Enzymol. Methods in Enzymology 65 (1): 201-11. ISBN 978-0-12-181965-1. PMID 6246343. doi:10.1016/S0076-6879(80)65028-9.
- Mishra, N. C.; Mishra, Nawin C. (1995). Molecular biology of nucleases. Boca Raton: CRC Press. pp. 46–52. ISBN 0-8493-7658-0.
- Douglas A. Julin (2000). «Detection and Quantitation of RecBCD Enzyme (Exonuclease V) Activity». DNA Repair Protocols. Methods in Molecular Biology 152. Humana Press. pp. 91-105. ISBN 978-0-89603-643-7. doi:10.1385/1-59259-068-3:91.
- Yang XC, Sullivan KD, Marzluff WF, Dominski Z (enero de 2009). «Studies of the 5′ Exonuclease and Endonuclease Activities of CPSF-73 in Histone Pre-mRNA Processing». Mol. Cell. Biol. 29 (1): 31-42. PMC 2612496. PMID 18955505. doi:10.1128/MCB.00776-08.
- West S, Gromak N, Proudfoot NJ (noviembre de 2004). «Human 5' → 3' exonuclease Xrn2 promotes transcription termination at co-transcriptional cleavage sites». Nature 432 (7016): 522-5. PMID 15565158. doi:10.1038/nature03035.
- Chen J, Chiang YC, Denis CL (marzo de 2002). «CCR4, a 3′–5′ poly(A) RNA and ssDNA exonuclease, is the catalytic component of the cytoplasmic deadenylase». EMBO J. 21 (6): 1414-26. PMC 125924. PMID 11889047. doi:10.1093/emboj/21.6.1414.
- Draper MP, Salvadore C, Denis CL (julio de 1995). «Identification of a mouse protein whose homolog in Saccharomyces cerevisiae is a component of the CCR4 transcriptional regulatory complex». Mol. Cell. Biol. 15 (7): 3487-95. PMC 230585. PMID 7791755.
- Moser MJ, Holley WR, Chatterjee A, Mian IS (diciembre de 1997). «The proofreading domain of Escherichia coli DNA polymerase I and other DNA and/or RNA exonuclease domains». Nucleic Acids Res. 25 (24): 5110-8. PMC 147149. PMID 9396823. doi:10.1093/nar/25.24.5110. Archivado desde el original el 18 de julio de 2012. Consultado el 7 de noviembre de 2009.
- Henry Y, Wood H, Morrissey JP, Petfalski E, Kearsey S, Tollervey D (mayo de 1994). «The 5' end of yeast 5.8S rRNA is generated by exonucleases from an upstream cleavage site». EMBO J. 13 (10): 2452-63. PMC 395111. PMID 7515008.
- Geerlings TH, Vos JC, Raué HA (diciembre de 2000). «The final step in the formation of 25S rRNA in Saccharomyces cerevisiae is performed by 5'→3' exonucleases». RNA 6 (12): 1698-703. PMC 1370040. PMID 11142370. doi:10.1017/S1355838200001540.
- Mukherjee D, et al. (2004). «Analysis of RNA Exonucleolytic Activities in Cellular Extracts». Springer protocols 257: 193-211. ISBN 1-59259-750-5. PMID 14770007. doi:10.1385/1-59259-750-5:193.
- Pamela A. Frischmeyer, et al. (2002). «An mRNA Surveillance Mechanism That Eliminates Transcripts Lacking Termination Codons». Science 295 (5563): 2258-61. PMID 11910109. doi:10.1126/science.1067338.
 Datos: Q411664
Datos: Q411664