Lactancia
El término lactancia describe la secreción de leche de las glándulas mamarias y el período de tiempo que una madre amamanta para alimentar a sus crías. El proceso ocurre naturalmente en todas las hembras de mamífero después del embarazo (véase embarazo humano), aunque su origen es anterior a los mamíferos.[1] En los seres humanos, el proceso de alimentación con leche también se llama lactancia materna o amamantamiento. Los bebés recién nacidos a menudo producen algo de leche de su propio tejido mamario, conocida coloquialmente como leche de bruja.


En la mayoría de las especies, la leche sale de los pezones de la madre; sin embargo, los monotremas, mamíferos que ponen huevos, carecen de pezones y liberan leche a través de conductos en el abdomen. En solo una especie de mamífero, el murciélago frugívoro Dayak del sudeste asiático, la producción de leche es una función masculina normal.
La galactopoyesis es el mantenimiento de la producción de leche. Esta etapa requiere prolactina. La oxitocina es fundamental para el reflejo de bajada de la leche en respuesta a la succión. La galactorrea es la producción de leche no relacionada con la lactancia. Puede ocurrir en machos y hembras de muchas especies de mamíferos como resultado de desequilibrios hormonales como la hiperprolactinemia.
Propósito
La función principal de la lactancia es proporcionar nutrición y protección inmunológica a las crías después del nacimiento. Debido a la lactancia, la pareja madre-cría puede sobrevivir incluso si la comida es escasa o demasiado difícil de alcanzar para las crías, lo que amplía las condiciones ambientales que la especie puede soportar. La costosa inversión de energía y recursos en la leche se ve compensada por el beneficio para la supervivencia de la descendencia.[2] En casi todos los mamíferos, la lactancia induce un período de infertilidad (en los seres humanos, amenorrea lactacional), que sirve para proporcionar el intervalo de nacimiento óptimo para la supervivencia de la descendencia.[3]
Humano
Influencias hormonales
A partir de la decimoctava semana de embarazo (segundo y tercer trimestre), el cuerpo de una mujer produce hormonas que estimulan el crecimiento del sistema de conductos lácteos en los senos:
- La progesterona influye en el crecimiento de tamaño de los alvéolos y lóbulos; los niveles altos de progesterona inhiben la lactancia antes del nacimiento. Los niveles de progesterona bajan después del nacimiento; esto desencadena el inicio de una abundante producción de leche.[4]
- El estrógeno estimula el crecimiento y la diferenciación del sistema de conductos lácteos. Al igual que la progesterona, los niveles altos de estrógeno también inhiben la lactancia. Los niveles de estrógeno también disminuyen en el momento del parto y permanecen bajos durante los primeros meses de lactancia. Las madres que amamantan deben evitar los métodos anticonceptivos basados en estrógenos, ya que un aumento en los niveles de estrógenos puede reducir el suministro de leche materna.
- La prolactina contribuye a un mayor crecimiento y diferenciación de los alvéolos, y también influye en la diferenciación de las estructuras ductales. Los altos niveles de prolactina durante el embarazo y la lactancia también aumentan la resistencia a la insulina, aumentan los niveles del factor de crecimiento (IGF-1) y modifican el metabolismo de los lípidos en preparación para la lactancia. Durante la lactancia, la prolactina es el factor principal que mantiene las uniones estrechas del epitelio ductal y regula la producción de leche a través del equilibrio osmótico.
- Lactógeno placentario humano (HPL): a partir del segundo mes de embarazo, la placenta libera grandes cantidades de HPL. Esta hormona está estrechamente asociada con la prolactina y parece ser fundamental en el crecimiento de la mama, el pezón y la areola antes del nacimiento.
- La hormona estimulante del folículo (FSH, del inglés Follicle stimulating hormone), la hormona luteinizante (LH) y la gonadotropina coriónica humana (hCG, del inglés human chorionic gonadotropin), mediante el control de la producción de estrógenos y progesterona, y también, por extensión, la producción de prolactina y hormona del crecimiento, son esenciales.
- La hormona del crecimiento (GH) es estructuralmente muy similar a la prolactina y contribuye independientemente a su galactopoyesis.
- La hormona adrenocorticotrópica (ACTH) y los glucocorticoides como el cortisol tienen una importante función inductora de la lactancia en varias especies animales, incluidos los seres humanos. Los glucocorticoides juegan un papel regulador complejo en el mantenimiento de uniones estrechas.
- La hormona estimulante de la tiroides (TSH) y la hormona liberadora de tirotropina (TRH) son hormonas galactopoyéticas muy importantes cuyos niveles aumentan naturalmente durante el embarazo.
- La oxitocina contrae el músculo liso del útero durante y después del nacimiento, y durante el orgasmo (s). Después del nacimiento, la oxitocina contrae la capa de músculo liso de células en forma de banda que rodean los alvéolos para exprimir la leche recién producida en el sistema de conductos. La oxitocina es necesaria para que se produzca el reflejo de eyección de la leche, o bajada, en respuesta a la succión.
También es posible inducir la lactancia sin embarazo. Los protocolos para inducir la lactancia se denominan protocolos Goldfarb. Usar píldoras anticonceptivas para imitar los niveles hormonales del embarazo, luego suspender el control de la natalidad, seguido del uso de un extractor de leche eléctrico doble durante sesiones de 15 minutos a intervalos regulares de 2-3 horas (más de 100 minutos en total por día) ayuda a inducir producción de leche.


Diferenciación secretora
Durante la última parte del embarazo, los senos de la mujer entran en la etapa de diferenciación secretora. Esto es cuando los senos producen calostro, un líquido espeso, a veces amarillento. En esta etapa, los niveles altos de progesterona inhiben la mayor parte de la producción de leche. No es una preocupación médica si una mujer embarazada pierde calostro antes del nacimiento de su bebé, ni es una indicación de la producción de leche en el futuro.
Activación secretora
Al nacer, los niveles de prolactina permanecen altos, mientras que el parto de la placenta resulta en una caída repentina de los niveles de progesterona, estrógeno y HPL. Esta abrupta retirada de progesterona en presencia de niveles altos de prolactina estimula la abundante producción de leche de activación secretora.
Cuando se estimula la mama, los niveles de prolactina en la sangre aumentan, alcanzan su punto máximo en unos 45 minutos y vuelven al estado anterior a la lactancia unas tres horas después. La liberación de prolactina hace que las células de los alvéolos produzcan leche. La prolactina también se transfiere a la leche materna. Algunas investigaciones indican que la prolactina en la leche es mayor en los momentos de mayor producción de leche y menor cuando los senos están más llenos, y que los niveles más altos tienden a ocurrir entre las 2 a. m. y las 6 a. m.[5]
Otras hormonas (en particular la insulina, la tiroxina y el cortisol) también están involucradas, pero sus funciones aún no se comprenden bien. Aunque los marcadores bioquímicos indican que la activación secretora comienza alrededor de 30 a 40 horas después del nacimiento, las madres no suelen comenzar a sentir un aumento de la plenitud de los senos (la sensación de que la leche "entra en el pecho") hasta 50 a 73 horas (2 a 3 días) después del nacimiento.
El calostro es la primera leche que recibe un bebé amamantado. Contiene cantidades más altas de glóbulos blancos y anticuerpos que la leche madura, y es especialmente alta en inmunoglobulina A (IgA), que recubre el revestimiento de los intestinos inmaduros del bebé y ayuda a evitar que los patógenos invadan el sistema del bebé. La IgA secretora también ayuda a prevenir las alergias alimentarias.[6] Durante las primeras dos semanas después del nacimiento, la producción de calostro da paso lentamente a la leche materna madura.[4]
Control autocrino - Galactapoyesis
El sistema de control endocrino hormonal impulsa la producción de leche durante el embarazo y los primeros días después del parto. Cuando el suministro de leche está más firmemente establecido, comienza el sistema de control autocrino (o local).
Durante esta etapa, cuanto más leche se extraiga de los senos, más leche producirá el seno.[7][8] La investigación también sugiere que drenar los senos más completamente también aumenta la tasa de producción de leche.[9] Por lo tanto, la producción de leche está fuertemente influenciada por la frecuencia con la que se alimenta el bebé y la capacidad de transferir la leche del pecho. El bajo suministro a menudo se puede atribuir a:
- no alimentarse o extraerse leche con la suficiente frecuencia
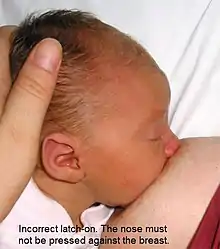
- incapacidad del bebé para transferir la leche de manera efectiva causada por, entre otras cosas:
- déficit en la estructura de la mandíbula o la boca
- mala técnica de enganche
- trastornos endocrinos maternos raros
- tejido mamario hipoplásico
- ingesta inadecuada de calorías o desnutrición de la madre
Reflejo de eyección de leche
Este es el mecanismo por el cual la leche se transporta desde los alvéolos mamarios hasta el pezón. La succión del bebé estimula los núcleos paraventriculares y el núcleo supraóptico en el hipotálamo, que envía señales a la glándula pituitaria posterior para que produzca oxitocina. La oxitocina estimula la contracción de las células mioepiteliales que rodean los alvéolos, que ya contienen leche. El aumento de presión hace que la leche fluya a través del sistema de conductos y se libere a través del pezón. Esta respuesta puede estar condicionada, por ejemplo, al llanto del bebé.
La expulsión de leche se inicia en el pecho de la madre por el acto de succionar por parte del bebé. El reflejo de expulsión de leche (también llamado reflejo de bajada) no siempre es constante, especialmente al principio. Una vez que una mujer está condicionada a amamantar, la bajada puede ser provocada por una variedad de estímulos, incluido el sonido de cualquier bebé. Incluso pensar en amamantar puede estimular este reflejo, provocando pérdidas no deseadas, o ambos senos pueden producir leche cuando un bebé se alimenta de un solo pecho. Sin embargo, este y otros problemas suelen resolverse después de dos semanas de alimentación. El estrés o la ansiedad pueden causar dificultades con la lactancia. La liberación de la hormona oxitocina conduce a la eyección de leche o al reflejo de bajada. La oxitocina estimula los músculos que rodean el seno para exprimir la leche. Las madres que amamantan describen la sensación de manera diferente. Algunos sienten un ligero cosquilleo, otros sienten una inmensa cantidad de presión o un ligero dolor/malestar, y otros no sienten nada diferente.
Un reflejo de eyección de leche deficiente puede deberse a pezones adoloridos o agrietados, separación del bebé, antecedentes de cirugía mamaria o daño tisular por traumatismo mamario anterior. Si una madre tiene problemas para amamantar, pueden ayudar diferentes métodos para ayudar al reflejo de expulsión de la leche. Estos incluyen amamantar en un lugar familiar y cómodo, masajear el pecho o la espalda o calentar el pecho con un paño o ducharse.
Mecanismo reflejo de eyección de leche
Este es el mecanismo por el cual la leche se transporta desde los alvéolos mamarios hasta el pezón. La succión del bebé inerva los mecanorreceptores de adaptación lenta[10] y de rápida adaptación[11] que están densamente agrupados alrededor de la región areolar. El impulso eléctrico sigue el tracto espinotalámico, que comienza con la inervación de los cuartos nervios intercostales. El impulso eléctrico luego asciende por el tracto posterolateral por uno o dos niveles vertebrales y hace sinapsis con neuronas de segundo orden, llamadas células del tracto, en el asta dorsal posterior. Las células del tracto luego se decusan a través de la comisura blanca anterior hacia la esquina anterolateral y ascienden al núcleo supraóptico y al núcleo paraventricular en el hipotálamo, donde hacen sinapsis con neuronas oxitocinérgicas de tercer orden. Los somas de estas neuronas se localizan en el hipotálamo, pero sus axones y terminales axónicos se localizan en el infundíbulo y la pars nerviosa de la hipófisis posterior, respectivamente. La oxitocina se produce en el soma de la neurona en los núcleos supraóptico y paraventricular, y luego se transporta hacia abajo por el infundíbulo a través del tracto hipotálamo-neurohipofisario con la ayuda de la proteína transportadora, neurofisina I, hasta la pars nerviosa de la hipófisis posterior y luego almacenados en cuerpos de Herring, donde se almacenan hasta la sinapsis entre neuronas de segundo y tercer orden.
Siguiendo el impulso eléctrico, la oxitocina se libera en el torrente sanguíneo. A través del torrente sanguíneo, la oxitocina llega a las células mioepiteliales, que se encuentran entre la matriz extracelular y las células epiteliales luminales que también forman los alvéolos en el tejido mamario. Cuando la oxitocina se une a las células mioepiteliales, las células se contraen. El aumento de la presión intraaveolar fuerza la leche hacia los senos lactíferos, hacia los conductos lactíferos (un estudio encontró que los senos lactíferos pueden no existir.[12] Si esto es cierto, la leche simplemente ingresa a los conductos lactíferos) y luego sale por el pezón.
Entuertos
Una oleada de oxitocina también hace que el útero se contraiga. Durante la lactancia, las madres pueden sentir estas contracciones como dolores posteriores. Estos pueden variar desde calambres menstruales hasta fuertes contracciones similares al trabajo de parto y pueden ser más graves con el segundo bebé y los siguientes.[13][14]
Sin embarazo, lactancia inducida, relactación
En los seres humanos, la lactancia y la relactación inducidas se han observado con frecuencia en algunas culturas y se han demostrado con éxito variable en las madres adoptivas. Parece plausible que la posibilidad de lactancia en mujeres (o hembras de otras especies) que no son madres biológicas confiera una ventaja evolutiva, especialmente en grupos con alta mortalidad materna y estrechos vínculos sociales.[15][16] El fenómeno también se ha observado en la mayoría de los primates, en algunos lémures y en mangostas enanas.[17][18]
La lactancia se puede inducir en seres humanos mediante una combinación de estimulación física y psicológica, mediante fármacos o mediante una combinación de esos métodos.[19] Algunas parejas pueden estimular la lactancia fuera del embarazo con fines sexuales.
Existen informes raros de la lactancia masculina (a diferencia de la galactorrea) en la literatura histórica médica y antropológica, aunque el fenómeno no ha sido confirmado por la literatura más reciente.[20]
La domperidona es un fármaco que puede inducir la lactancia.[21][22]
Evolución
Charles Darwin reconoció que las glándulas mamarias parecían haberse desarrollado específicamente a partir de glándulas cutáneas, y planteó la hipótesis de que evolucionaron a partir de glándulas en bolsas de cría de peces, donde proporcionarían alimento a los huevos.[1] El último aspecto de su hipótesis no ha sido confirmado; sin embargo, más recientemente se ha postulado el mismo mecanismo para los primeros sinápsidos.[23]
Como todos los mamíferos lactan, la lactancia debe haber evolucionado antes que el último ancestro común de todos los mamíferos, lo que la sitúa en un mínimo en el Triásico Medio o Tardío cuando los monotremas divergieron de los terios.[24] O. T. Oftedal ha argumentado que los terápsidos desarrollaron un fluido proto-lácteo para mantener los huevos húmedos, una adaptación necesaria debido a los huevos con cáscara de pergamino de diápsidos que son más vulnerables a la evaporación y deshidratación que los huevos mineralizados producidos por algunos saurópsidos.[23][25] Este fluido protolacteo se convirtió en una leche compleja y rica en nutrientes que luego permitió una disminución en el tamaño del huevo al reducir la dependencia de una yema grande en el huevo. También se cree que la evolución de la lactancia dio lugar a una dentición más compleja que se observa en los mamíferos, ya que la lactancia habría permitido el desarrollo prolongado de la mandíbula antes de la erupción de los dientes.
Durante la evolución temprana de la lactancia, la secreción de leche era a través de glándulas pilosebáceas en parches mamarios, análogos a la areola, y los pelos en este parche transportaban los fluidos nutritivos a las crías como se ve en los monotremas.[26] El desarrollo del pezón ocurrió en los linajes de mamíferos que divergieron después de los monotremas, metatheria y eutheria.
Ocurrencia fuera de Mammalia
Otro ejemplo bien conocido de cría nutritiva con secreciones de glándulas es la leche de buche de aves columbiformes. Como en los mamíferos, esto también parece estar dirigido por la prolactina.[27] Otras aves, como los flamencos y los pingüinos, utilizan técnicas de alimentación similares.[28]
El pez disco (Symphysodon) es conocido por alimentar (biparentalmente) a sus crías mediante la secreción de moco epidérmico.[29][30] Un examen más detenido revela que, como en los mamíferos y las aves, la secreción de este fluido nutritivo puede ser controlada por la prolactina.[31] Se observa un comportamiento similar en al menos 30 especies de cíclidos.
La lactancia es también el sello distintivo de la viviparidad adenotrófica, un mecanismo de reproducción desarrollado por algunos insectos, sobre todo las moscas tsetsé. El único óvulo de la mosca tsetsé se convierte en una larva dentro del útero, donde es alimentado por una sustancia lechosa secretada por una glándula mamaria dentro del útero.[32] También se sabe que la especie de cucaracha Diploptera punctata alimenta a sus crías con secreciones lechosas.[33]
Toxeus magnus, una especie de araña saltarina que imita a las hormigas del sudeste asiático, también lacta. Amamanta a su descendencia durante unos 38 días, aunque pueden alimentarse por sí mismos después de 21 días. El bloqueo de la lactancia inmediatamente después del nacimiento resultó en la mortalidad completa de la descendencia, mientras que el bloqueo 20 días después del nacimiento resultó en un aumento de la búsqueda de alimento y una reducción de la supervivencia. Esta forma de lactancia puede haber evolucionado a partir de la producción de huevos tróficos.[34]
Véase también
- Sala de lactancia
- Galactogogo
- Línea de leche
- Lactancia masculina
- Ubre
- Amamantamiento
- Fracaso de la lactancia
- Supresión de la lactancia
- Lactancia erótica
- Eje hipotalámico-pituitario-prolactina
- Caritas romana
- El embarazo
Referencias
- Capuco, A. V.; Akers, R. M. (2009). «The origin and evolution of lactation». Journal of Biology 8 (4): 37. PMC 2688910. PMID 19439024. doi:10.1186/jbiol139.
- Power, Michael L. (2016). Milk : the biology of lactation. ISBN 978-1-4214-2043-1. OCLC 954481825.
- McNeilly, A. S. (1997-07-XX). «Lactation and fertility». Journal of Mammary Gland Biology and Neoplasia 2 (3): 291-298. ISSN 1083-3021. PMID 10882312. doi:10.1023/a:1026340606252.
- Mohrbacher, Nancy; La Leche League International (2003). The breastfeeding answer book (3rd rev. ed edición). La Leche League International. ISBN 0-912500-92-1. OCLC 51844325.
- Cregan, Mark D.; Mitoulas, Leon R.; Hartmann, Peter E. (2002-03). «Milk prolactin, feed volume and duration between feeds in women breastfeeding their full-term infants over a 24 h period». Experimental Physiology 87 (2): 207-214. ISSN 0958-0670. PMID 11856965. doi:10.1113/eph8702327.
- Sears, Martha (2000). The breastfeeding book : everything you need to know about nursing your child from birth through weaning (1st ed edición). Little Brown. ISBN 0-316-77727-7. OCLC 42849740.
- deCarvalho, M.; Anderson, D. M.; Giangreco, A.; Pittard, W. B. (1985-05-XX). «Frequency of milk expression and milk production by mothers of nonnursing premature neonates». American Journal of Diseases of Children (1960) 139 (5): 483-485. ISSN 0002-922X. PMID 3984973. doi:10.1001/archpedi.1985.02140070057033.
- Hopkinson, J. M.; Schanler, R. J.; Garza, C. (1988-06-XX). «Milk production by mothers of premature infants». Pediatrics 81 (6): 815-820. ISSN 0031-4005. PMID 3368280.
- Daly S, Owens R, Hartmann P; Owens; Hartmann (1993). «The short-term synthesis and infant-regulated removal of milk in lactating women». Exp Physiol 78 (2): 209-20. PMID 8471241. doi:10.1113/expphysiol.1993.sp003681.
- Grachev, I. I.; Alekseev, N. P.; Velling, V. A. (1977-03-XX). «[Slowly-adapting mechanoreceptor units of the guinea pig mammary nipple]». Fiziologicheskii zhurnal SSSR imeni I. M. Sechenova 63 (3): 391-400. ISSN 0015-329X. PMID 863036.
- Grachev, I. I.; Alekseev, N. P.; Velling, V. A. (1976). «[Properties of the mechanoreceptors of the nipple of the guinea pig mammary gland. (Rapidly adapting mechanoreceptor units)]». Fiziologicheskii zhurnal SSSR imeni I. M. Sechenova 62 (6): 885-892. ISSN 0015-329X. PMID 1010088.
- Ramsey, DT; Kent, JC; Hartmann, RA; Hartmann, PE (2005). «Anatomy of the lactating human breast redefined with ultrasound imaging». Journal of Anatomy 206 (6): 525-34. PMC 1571528. PMID 15960763. doi:10.1111/j.1469-7580.2005.00417.x.
- Breastfeeding Answers Made Simple, Nancy Mohrbacher, IBCLC, FILCA
- Fray, Kathy (2005). Oh baby-- : birth, babies & motherhood uncensored. Random House. ISBN 1-86941-713-5. OCLC 156774948.
- Sobrinho, Luis Gonçalves (2003). «Prolactin, psychological stress and environment in humans: adaptation and maladaptation». Pituitary 6 (1): 35-39. ISSN 1386-341X. PMID 14674722. doi:10.1023/a:1026229810876.
- Bose, C. L.; D'Ercole, A. J.; Lester, A. G.; Hunter, R. S.; Barrett, J. R. (1981-04-XX). «Relactation by mothers of sick and premature infants». Pediatrics 67 (4): 565-569. ISSN 0031-4005. PMID 6789296.
- König, B. (1997). «Cooperative care of young in mammals». Die Naturwissenschaften 84 (3): 95-104. Bibcode:1997NW.....84...95K. PMID 9112240. doi:10.1007/s001140050356.
- Creel, S. R.; Monfort, S. L.; Wildt, D. E.; Waser, P. M. (1991). «Spontaneous lactation is an adaptive result of pseudopregnancy». Nature 351 (6328): 660-662. Bibcode:1991Natur.351..660C. PMID 2052092. doi:10.1038/351660a0.
- Seema; Patwari, AK; Satyanarayana, L (1997). «Relactation: an effective intervention to promote exclusive breastfeeding». Journal of Tropical Pediatrics 43 (4): 213-6. PMID 9283123. doi:10.1093/tropej/43.4.213.
- Strange but True: Males Can Lactate: Scientific American
- https://www.theguardian.com/science/2018/feb/14/transgender-woman-breastfeed-health
- https://www.liebertpub.com/doi/10.1089/trgh.2017.0044
- Oftedal, Olav T. (2002-07). «The mammary gland and its origin during synapsid evolution». Journal of Mammary Gland Biology and Neoplasia 7 (3): 225-252. ISSN 1083-3021. PMID 12751889. doi:10.1023/a:1022896515287.
- van Rheede, Teun; Bastiaans, Trijntje; Boone, David N.; Hedges, S. Blair; de Jong, Wilfried W.; Madsen, Ole (2006-03). «The platypus is in its place: nuclear genes and indels confirm the sister group relation of monotremes and Therians». Molecular Biology and Evolution 23 (3): 587-597. ISSN 0737-4038. PMID 16291999. doi:10.1093/molbev/msj064.
- Oftedal, Olav T. (2002-07). «The origin of lactation as a water source for parchment-shelled eggs». Journal of Mammary Gland Biology and Neoplasia 7 (3): 253-266. ISSN 1083-3021. PMID 12751890. doi:10.1023/a:1022848632125.
- Oftedal, Olav T.; Dhouailly, Danielle (2013-06). «Evo-devo of the mammary gland». Journal of Mammary Gland Biology and Neoplasia 18 (2): 105-120. ISSN 1573-7039. PMID 23681303. doi:10.1007/s10911-013-9290-8.
- Horseman, N. D.; Buntin, J. D. (1995). «Regulation of pigeon cropmilk secretion and parental behaviors by prolactin». Annual Review of Nutrition 15: 213-238. ISSN 0199-9885. PMID 8527218. doi:10.1146/annurev.nu.15.070195.001241.
- «Bird Milk». web.stanford.edu. Consultado el 8 de abril de 2021.
- Buckley, Jonathan; Maunder, Richard J.; Foey, Andrew; Pearce, Janet; Val, Adalberto L.; Sloman, Katherine A. (15 de noviembre de 2010). «Biparental mucus feeding: a unique example of parental care in an Amazonian cichlid». The Journal of Experimental Biology 213 (Pt 22): 3787-3795. ISSN 1477-9145. PMID 21037057. doi:10.1242/jeb.042929.
- Chong, Kenny; Joshi, Shashikant; Jin, Lam Toong; Shu-Chien, Alexander Chong (2006-04). «Proteomics profiling of epidermal mucus secretion of a cichlid (Symphysodon aequifasciata) demonstrating parental care behavior». Proteomics 6 (7): 2251-2258. ISSN 1615-9853. PMID 16385477. doi:10.1002/pmic.200500591.
- Khong, Hou-Keat; Kuah, Meng-Kiat; Jaya-Ram, Annette; Shu-Chien, Alexander Chong (2009-05). «Prolactin receptor mRNA is upregulated in discus fish (Symphysodon aequifasciata) skin during parental phase». Comparative Biochemistry and Physiology. Part B, Biochemistry & Molecular Biology 153 (1): 18-28. ISSN 1879-1107. PMID 19272315. doi:10.1016/j.cbpb.2009.01.005.
- Attardo, G. M.; Lohs, C.; Heddi, A.; Alam, U. H.; Yildirim, S.; Aksoy, S. (2008). «Analysis of milk gland structure and function in Glossina morsitans: Milk protein production, symbiont populations and fecundity». Journal of Insect Physiology 54 (8): 1236-1242. PMC 2613686. PMID 18647605. doi:10.1016/j.jinsphys.2008.06.008.
- Williford, Anna; Stay, Barbara; Bhattacharya, Debashish (2004-03-XX). «Evolution of a novel function: nutritive milk in the viviparous cockroach, Diploptera punctata». Evolution & Development 6 (2): 67-77. ISSN 1520-541X. PMID 15009119. doi:10.1111/j.1525-142x.2004.04012.x.
- Chen, Zhanqi; Corlett, Richard T.; Jiao, Xiaoguo; Liu, Sheng-Jie; Charles-Dominique, Tristan; Zhang, Shichang; Li, Huan; Lai, Ren et al. (30 de noviembre de 2018). «Prolonged milk provisioning in a jumping spider». Science (New York, N.Y.) 362 (6418): 1052-1055. ISSN 1095-9203. PMID 30498127. doi:10.1126/science.aat3692.
Enlaces externos
- ¿Cómo perdieron los mamíferos sus yemas de huevo? ¿Los mamíferos desarrollaron leche nutritiva antes o después de abandonar los huevos con yemas? (New Scientist, 18 de marzo de 2008)