Maraapunisaurus fragillimus
Maraapunisaurus fragillimus (del Ute "lagarto enorme muy frágil") es la única especie conocida del género extinto Maraapunisaurus de dinosaurios saurópodo rebaquisáurido, que vivió a finales del Jurásico, hace aproximadamente 150 y 147 millones de años, en el Kimmeridgiense y el Titoniense, en lo que hoy es Norteamérica.[2]
| Maraapunisaurus fragillimus | ||
|---|---|---|
| Rango temporal: 150,8 Ma - 145,5 Ma [1] Jurásico Superior | ||
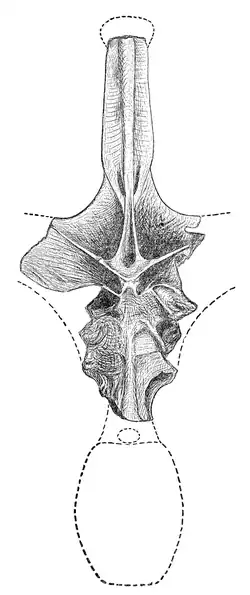 Ilustración de Cope del espécimen de Maraapunisaurus fragillimus | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Sauropoda | |
| Familia: | Rebbachisauridae | |
| Género: |
Maraapunisaurus Carpenter, 2018 | |
| Especie: |
M. fragillimus Cope, 1878 | |
| Sinonimia | ||
| ||
Descripción
Para tener una estimación del tamaño completo del M. fragillimus, se usó escalas con huesos conocidos de diplodócidos, una familia de saurópodos extremadamente largos y delgados, siempre del que sus proporciones relativas fueron similares. En su trabajo original, Cope hizo esto, especulando acerca del tamaño del fémur, hueso superior de la pierna, hipotético de M. fragillimus. Cope notó que en otros dinosaurios saurópodos, específicamente en Amphicoelias altus y en Camarasaurus supremus, el fémur era siempre dos veces más alto que la vértebra dorsal más alta, y estimaba el tamaño de un fémur del M fragillimus en 3,6 metros de altura.[3] En 1994, usando los diplodocos como referencia, Gregory S. Paul estimó una longitud del fémur de 3,1 a 4 metros para el M. fragillimus.[4] La nueva evaluación de 2006 del M. fragillimus de Ken Carpenter también utilizó los diplodocos como guía de la escala, encontrando una altura del fémur de 4,3 a 4,6 metros. Carpenter comenzó a estimar el tamaño completo de M. fragillimus, aunque advirtió que las proporciones relativas en diplodócidos podrían variar entre especies. Si se asume que las proporciones son las mismas de los Diplodocus, Carpenter presentó una longitud total estimada de 58 metros, estimación que está dentro de la gama presentada por Paul en 1994, que fue de 40 a 60 metros.[4] Carpenter precisó que incluso las estimaciones más bajas de la longitud para M. fragillimus eran más altas que las de los otros saurópodos gigantes, tales como el diplodócido Supersaurus de 32,5 metros, el braquiosáurido Sauroposeidon de 30 metros y el titanosauriano Argentinosaurus de 39,7 metros.[5][6]
Carpenter presentó proporciones más especulativas y específicas para M. fragillimus, otra vez, basadas en Diplodocus, incluyendo una longitud del cuello de 16,75 metros, una longitud de cuerpo de 9,25 metros y una longitud de la cola de 32 metros. Él estimaba la altura total de la pata delantera en 5,75 metros y la altura del miembro trasero en 7,5 metros, y la altura total, en el punto más alto de la espalda en 9,25 metros. La ballena azul, considerada como el vertebrado vivo más largo, alcanza entre 30 a 33 metros de longitud.[5][7]
Mientras que M. fragillimus era relativamente delgado, su tamaño enorme lo hizo muy masivo. El peso es mucho más difícil de determinar que la longitud en saurópodos, pues las ecuaciones necesarias, más complejas, son propensas a mayores márgenes de error basados en variaciones más pequeñas en las proporciones totales del animal. Carpenter utilizó la estimación de Paul de 1994 de la masa del Diplodocus carnegii de 11,5 toneladas para especular que M. fragillimus habría podido pesar hasta 122,4 toneladas, al principio se estimó como máximo 185 toneladas, actualmente descartado.[5] La ballena azul más pesada en el expediente pesaba cerca de 173 toneladas,[8] y el dinosaurio más pesado conocido por restos razonablemente buenos, el Argentinosaurus, de 83,2 toneladas,[6] aunque si las estimaciones del tamaño pueden ser validadas, todavía sería más ligero que Bruhathkayosaurus, del que se estima que pudo pesar entre 126 y 157 toneladas, con base en una descripción de un fósil muy parcial, también muy cuestionado.[9][10] E inicialmente se le había estimado con un peso de 175-220 toneladas (estimación que posteriormente fue renegado por el autor).[11]
Estudios más recientes han hecho una serie de sugerencias con respecto a la posibilidad de tal animal. Un análisis de la evidencia sobreviviente y la plausibilidad biológica de un animal terrestre tan grande, ha sugerido que el enorme tamaño de este animal era sobreestimaciones debidas en parte a errores tipográficos en la descripción original de 1878.[12] Más recientemente, el paleontólogo Kenneth Carpenter sugirió que la especie era un saurópodo rebaquisáurido y no un diplodócido, y por lo tanto usó a Limaysaurus en lugar de Diplodocus como base para las estimaciones de tamaño. Esto resultó en un animal más pequeño, de 30,3 metros de largo y 7,95 metros en las caderas, y descartó la idea de que debió haber errores tipográficos.[2] Sin embargo, Parrish en 2006 encontró que la longitud del cuello en saurópodos escalado al largo del torso a un índice de 1.35.[13] Esto significaría que el cuello de Maraapunisaurus fue de 7,6 metros de largo en lugar de los 6 metros de largo, que tendría si Limaysaurus simplemente se escalara más grande. El resultado es que Maraapunisaurus podría haber sido de 32 metros de largo, lo que lo coloca dentro del rango estimado para Supersaurus y Diplodocus hallorum.[14][2]
Descubrimiento e investigación
Maraapunisaurus está presente en la zona estratigráfica 6 de la Formación Morrison.[15] Originalmente llamado Amphicoelias fragillimus que debido al extravío de los restos es considerada dudosa.[16] Basados en la descripción del posible único fósil de un M. fragillimus, que significa "muy frágil", en referencia a la dureza de sus huesos, este debió haber sido el más largo de los vertebrados terrestres, con entre 40 y 60 metros de largo y una masa de alrededor de 122 toneladas, rivalizando con el animal más grande conocido, la ballena azul. Sin embargo, debido a que los huesos se han perdido y los estudios se realizaron en la década de 1870, sólo quedan los dibujos del cuaderno de campo de quien lo describió.
El holotipo y único espécimen conocido de Maraapunisaurus fragillimus fue recolectado por Oramel Lucas, poco después de haber sido contratado como coleccionista de fósiles empleado por el reconocido paleontólogo Edward Drinker Cope, en 1877. Lucas descubrió una vértebra parcial en Garden Park, al norte de Cañon City, Colorado, cerca de la cantera que produjo los primeros especímenes de Camarasaurus. La vértebra estaba en mal estado, pero era sorprendentemente grande, midiendo entre 1,5 metros hasta 2.7 metros de altura.[5]
Lucas envió el espécimen a comienzos del verano boreal de 1878, y Cope lo publicó como el espécimen holotipo (número de catálogo AMNH 5777) de una nueva especie, M. fragillimus. El nombre deriva del griego fragillimus, que significa "muy frágil", se refiere a la fragilidad del hueso, producida por sus láminas muy finas de las paredes vertebrales. Según revelan los cuadernos de Cope, que él registró con base en el informe de Lucas sobre la localización del sitio de la excavación en 1879, el espécimen vino del sur de la colina de la misma mina que Camarasaurus, ahora conocida como "Cope's Nipple".[3] Aunque Cope originalmente escribió que el sitio pertenecía a la formación Dakota de mediados del Cretácico, pertenecen a la formación Morrison, que sitúa la edad del sitio en 150 millones de años en el período Jurásico Superior, específicamente el Titoniense.[17]
El gigante desmenuzado
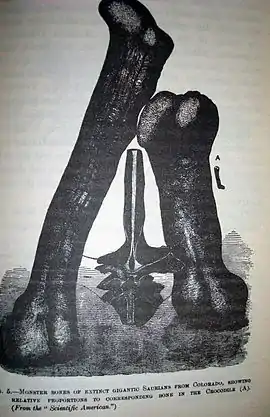
A los huesos gigantescos atribuidos a M. fragillimus no se les ha hecho caso muy a menudo en los resúmenes de los dinosaurios más grandes, en parte porque, según varios informes subsecuentes, el paradero de la vértebra y el fémur es desconocido, y todas las tentativas de localizarlos han fallado. Carpenter, en 2006, presentó una teoría posible para explicar la desaparición de los especímenes de M. fragillimus. Cope observó en su descripción que el material de los arcos neurales era muy frágil, y las técnicas para endurecer y preservar el hueso fósil todavía no habían sido inventadas (el rival de Cope, el paleontólogo O. C. Marsh, fue el primero en utilizar tales productos químicos, en la década de 1880). Carpenter observó que los huesos fósiles conocidos de la mina de M. fragillimus habían sido preservados en lodolita, la cual tiende a desmenuzarse fácilmente en fragmentos en forma de cubos pequeños e irregulares. Por lo tanto, el espécimen pudo haberse desmenuzado y desechado por Cope poco después de que él lo ilustrara para su trabajo (Carpenter sugirió que esto puede explicar por qué Cope dibujó la vértebra en solamente una vista, en lugar de desde ángulos múltiples, como hizo con sus otros descubrimientos).[5]
En 1994, fue hecha una tentativa de volver a la mina original donde M. fragillimus y otras especies habían sido encontradas, usando un radar de penetración terrestre para localizar huesos, cuya imagen debería mostrarlos enterrados en la tierra. Esta tentativa falló, debido al hecho de que los huesos fosilizados en lodolita eran de la misma densidad que la roca circundante, haciéndolos imposibles de distinguir. Un estudio de la topografía local también demostró que los estratos de la roca del fósil fueron seriamente erosionados, y estaban probablemente así cuando Lucas hizo su descubrimiento de M. fragillimus, indicando que la mayoría del esqueleto pudo ya estar perdida para el momento en que la vértebra y el fémur fueron recuperados.
Carpenter en 2006 también observó que, debido al tamaño extraordinario y a la desaparición misteriosa del fósil, la descripción de Cope de M. fragillimus ha sido tomada con escepticismo por alguna gente que asume que había errores tipográficos en sus medidas. Carpenter discutió que había razones para fiarse de la palabra de Cope, observando que la reputación de los paleontólogos estaba en juego. El descubrimiento ocurrió durante las Guerra de los Huesos y Marsh, su rival, quien estaba "siempre listo para humillar" a Cope, nunca puso en duda sus afirmaciones acerca de Amphicoelias. Marsh era conocido por tener espías empleados en supervisar los descubrimientos de Cope, y pudo incluso haber tenido confirmación del tamaño enorme de los huesos. Osborn y Mook en 1921 y McIntosh en 1998 también aceptaron los datos de Cope sin discutirlos.[5][18][19] Otros paleontólogos han sido más críticos. En un análisis de la evidencia y las circunstancias que rodearon la publicación e interpretación del descubrimiento, Woodruff y Foster de 2015 concluyeron que su tamaño había sido sobrestimado y que los paleontólogos modernos estaban aceptando la interpretación de Cope sin el debido escepticismo. Señalan que no se han descubierto fósiles de saurópodos semejantemente gigantescos en la Formación Morrison ni en ninguna otra parte, que los paleontólogos del siglo XIX, incluido el propio Cope, no prestaron atención al tamaño de los fósiles, incluso cuando puede haber comprobado el aumento de tamaño de la Regla de Cope en linajes animales a lo largo del tiempo, y que los errores tipográficos en sus mediciones, como el informe de mediciones vertebrales en metros en lugar de milímetros, socavan su fiabilidad. Llegaron a la conclusión de que el supergigante M. fragillimus es una criatura "altamente improbable" basada en una interpretación incondicional del informe de Cope.[12]
En 2018, Carpenter defendió una vez más las medidas originales dadas por Cope en un documento que redacta las especies basándose en la ilustración y descripción de Cope. El argumento de un error tipográfico apuntaba al uso de Cope de la abreviatura "m" en lugar de "mm" para los milímetros, el mismo que se usó para los metros, pero Carpenter señala que se utiliza una "M" mayúscula para los metros, y que esto fue un método estándar de abreviatura en esa época. Además de esto, señaló la comunicación entre Lucas y Ferdinand Vandeveer Hayden, un geólogo, donde se repitió el gran tamaño sin cuestionarlo. Lucas también hizo sus propios dibujos anotados con medidas más específicas que las dadas por Cope, por lo que Carpenter considera que es poco probable que simplemente estuviera replanteando lo que Cope había dicho. Más tarde, en 1880, Lucas incluyó una mención específica del espécimen en su autobiografía, señalando "Qué monstruo debe haber sido el animal", refutando la idea de que no se prestó atención a la importancia de la vértebra. Carpenter fue crítico de la teoría del error tipográfico, diciendo: "Es desafortunado que busquen desacreditar la afirmación de Cope al lanzar calumnias acerca de la calidad de su trabajo e ir tan lejos como alterar las medidas dadas por él".[2]
Al estudiar nuevamente el dibujo del espécimen, Carpenter descubrió que mostraba una gran semejanza con la vértebra de los diplodocoides rebaquisáuridos. Anteriormente se lo había relacionado con la especie tipo de Amphicoelias, A. altus, como un diplodócido o un diplodocoide primitivo. Al considerar que se trataba de un rebaquisáurido en su reanálisis, la especie no podía ser referida al género Amphicoelias, por lo que le dio un nuevo nombre genérico, Maraapunisaurus. Aparentemente Carpenter fue inspirado por el trabajo de entusiastas de la paleontología publicado en el sitio web DeviantArt; uno de sus usuarios, Zachary Armstrong, conocido allí como "palaeozoologist", predijo esta clasificación correctamente en 2014, cuatro años antes de que Carpenter la confirmara. Armstrong fue reconocido en los agradecimientos del artículo científico. Nombrar un género basado en un espécimen perdido es algo raro, pero señaló que el ICZN no lo prohíbe, y que el género Nopcsaspondylus fue nombrado de manera similar. El nombre del género se deriva del término en ute sureño "Ma-ra-pu-ni", que significa "grande", y el griego latinizado saurus, que significa lagarto. El nombre fue sugerido a Carpenter por el Departamento Cultural de los Ute del Sur, con base en Ignacio (Colorado).[2]
Clasificación
En 1921, Osborn y Mook colocaron a A. fragillimus en la familia Diplodocidae.[20] Esto fue generalmente aceptado hasta bien entrado el siglo XXI. En 2018, Carpenter concluyó de una comparación anatómica cualitativa que Maraapunisaurus era un miembro basal de los Rebbachisauridae. La posición basal estaría indicada por la presencia de un hiposfeno, un proceso secundario de articulación posterior, que es un rasgo compartido con los rebaquisáuridos basales Histriasaurus, Comahuesaurus y Demandasaurus. Maraapunisaurus podría ser más derivado que Histriasaurus se muestra por la extensa neumatización y la alta base del arco neural.[2]
Si Maraapunisaurus pertenece al Rebbachisauridae, sería el miembro más antiguo de su grupo conocido y el único descubierto del Jurásico. El anterior más antiguo fue Xenoposeidon. También sería el único de América del Norte. Carpenter concluyó que el Rebbachisauridae podría haberse originado en ese continente y solo más tarde se extendió a Europa, desde allí habrían invadido América del Sur. La interpretación habitual había sido que los rebbachisauridos eran de origen sudamericano.[2]
Paleoecología

En su revaluación de 2006, Carpenter examinó la paleobiología de los saurópodos gigantes, incluido Maraapunisaurus, y abordó la cuestión de por qué este grupo alcanzó un tamaño tan grande. Señaló que los tamaños gigantescos se alcanzaron temprano en la evolución de los saurópodos, con especies de tamaño muy grande presentes tan temprano como en el Triásico tardío, y concluyó que cualquier presión evolutiva causada por el gran tamaño estaba presente desde los orígenes tempranos del grupo. Carpenter citó varios estudios de herbívoros gigantes de mamíferos, como elefantes y rinocerontes, que mostraron que el mayor tamaño en animales que comen plantas conduce a una mayor eficiencia en la digestión de alimentos. Dado que los animales más grandes tienen sistemas digestivos más largos los alimentos se mantienen en la digestión durante períodos de tiempo significativamente más largos, lo que permite que los animales grandes sobrevivan con fuentes de alimentos de menor calidad. Esto es especialmente cierto en animales con un gran número de "cámaras de fermentación" a lo largo del intestino, que permiten que los microbios se acumulen y fermenten el material vegetal, ayudando a la digestión. A lo largo de su historia evolutiva, los dinosaurios saurópodos se encontraron principalmente en ambientes semiáridos, estacionalmente secos, con una correspondiente caída estacional en la calidad de los alimentos durante la estación seca. El ambiente de Maraapunisaurus era esencialmente una sabana, similar a los ambientes áridos en los que se encuentran los herbívoros gigantes modernos, apoyando la idea de que los alimentos de baja calidad en un ambiente árido promueven la evolución de los herbívoros gigantes. Carpenter argumentó que otros beneficios del gran tamaño, como la inmunidad relativa de los depredadores, un menor gasto de energía y una mayor vida útil, son probablemente ventajas secundarias.[5]
El entorno de la Formación Morrison en el que vivía Maraapunisaurus se habría parecido a una sabana moderna, aunque como el pasto no apareció hasta el Cretácico Superior, los helechos fueron probablemente la planta dominante y la principal fuente de alimento para Maraapunisaurus. Aunque Engelmann et al. en 2004 rechazaron los helechos como fuente de alimento para los saurópodos debido a su contenido calórico relativamente bajo,[21] Carpenter argumentó que el sistema digestivo saurópodo, bien adaptado para manejar alimentos de baja calidad, permite el consumo de helechos como una gran parte de la dieta de estos animales.[5] Carpenter también observó que la presencia ocasional de grandes troncos petrificados indica la presencia de árboles de 20 a 30 metros de altura, lo que parece estar en conflicto con la comparación de la sabana. Sin embargo, los árboles son raros, y dado que los árboles altos requieren más agua de la que el ambiente de la sabana podría proporcionar, probablemente existieron en zonas estrechas o "bosques de galería" a lo largo de los ríos y barrancos donde podría acumularse agua. Carpenter especuló que herbívoros gigantes como Maraapunisaurus pueden haber usado la sombra de los bosques de la galería para mantenerse frescos durante el día, y haberse alimentado en la sabana en la mayor parte de la noche.[5]
Referencias
- «Amphicoelias». Paleobiology Database (en inglés).
- Carpenter, Kenneth (2018). "Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado". Geology of the Intermountain West. 5: 227–244.
- Cope, E.D. (1878a). "On the Vertebrata of the Dakota Epoch of Colorado." Proceedings of the American Philosophical Society, 17: 233-247.
- Paul, G.S. (1994a). "Big sauropods - really, really big sauropods." The Dinosaur Report, The Dinosaur Society, Fall, p. 12-13.
- Carpenter, Kenneth (2006). Foster, John R.; Lucas, Spencer G., ed. «Biggest of the Big: A Critical Re-Evaluation of the Mega-Sauropod Amphicoelias fragillimus Cope, 1878» 36. New Mexico Museum of Natural History and Science Bulletin. pp. 131-138. Archivado desde el original el 8 de marzo de 2016. Consultado el 22 de octubre de 2018.
- Sellers, William; Margetts, Lee; Coria, Rodolfo; Manning, Phillip (30 de octubre de 2013). March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs (en inglés). PMID 24348896. doi:10.1371/journal.pone.0078733. Consultado el 11 de septiembre de 2015.
- J. Calambokidis and G. Steiger (1998). Blue Whales. Voyageur Press. ISBN 0-89658-338-4.
- «Assessment and Update Status Report on the Blue Whale Balaenoptera musculus» (PDF) (en inglés). Committee on the Status of Endangered Wildlife in Canada. 2002. Consultado el 11 de septiembre de 2015.
- Wedel, Matt (20 de mayo de 2008). «SV-POW! showdown: sauropods vs whales» (en inglés). Consultado el 11 de septiembre de 2015.
- Mortimer, Mickey (7 de septiembre de 2004). «Re: Largest Dinosaurs» (en inglés). Archivado desde el original el 13 de septiembre de 2019. Consultado el 11 de septiembre de 2015.
- Mortimer, Mickey (19 de junio de 2001). «Re: Bruhathkayosaurus» (en inglés). Archivado desde el original el 22 de mayo de 2017. Consultado el 11 de septiembre de 2015.
- Woodruff, C; Foster, JR (2015). «The fragile legacy of Amphicoelias fragillimus (Dinosauria: Sauropoda; Morrison Formation - Latest Jurassic)». PeerJ PrePrints. doi:10.7287/peerj.preprints.838v1.
- Parrish, J.M., 2006, The origins of high browsing and the effects of phylogeny and scaling on neck length in sauropodomorphs, in Carrano, M.T., Gaudin, T.J., Blob, R.W., and Wible, J.R., editors, Amniote paleob
- Lovelace, D.M., Hartman S.A., and Wahl, W.R., 2008, Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny: Arquivos do Museu Nacional, Rio de Janeiro, v. 65, no. 4, p. 527–544.
- Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
- HE NRY GALIANO ¹ AND RAIMU ND ALBERSDÖRFER; THE MORRISON FORMATION, BIG HORN BASIN, WYOMING, WITH TAXONOMIC REEVALUATION OF DIPLODOCUS, APATOSAURUS, BAROSAURUS, AND OTHER GENERA Archivado el 13 de marzo de 2012 en Wayback Machine.; 2010 Dinosauria International, LLC, Wyoming: ISBN 978-0-9830585-0-2
- Turner, C.E., and Peterson, F. (1999) "Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A." In. Gillette, D., ed., Vertebrate Paleontology in Utah: Utah Geological Survey Miscellaneous Publication, 99(1): 77–114.
- Osborn, H.F., and Mook, C. C. (1921). "Camarasaurus, Amphicoelias and other sauropods of Cope." Memoirs of the American Museum of Natural History NS, 3(3): 249-387.
- McIntosh, J.S. (1998) "New information about the Cope collection of sauropods from Garden Park, Colorado." In Carpenter, K., Chure, D. and Kirkland, J.I., eds., The Morrison Formation: an interdisciplinary study: Modern Geology, 23: 481-506.
- Osborn, H.F.; Mook, C. C. (1921). «Camarasaurus, Amphicoelias and other sauropods of Cope». Memoirs of the American Museum of Natural History NS 3 (3): 249-387.
- Engelmann, G.F., Chure, D.J., and Fiorillo, A.R. (2004). "The implications of a dry climate for the paleoecology of the fauna of the Upper Jurassic Morrison Formation." In Turner, C.E., Peterson, F., and Dunagan, S.P., eds., Reconstruction of the extinct ecosystem of the Upper Jurassic Morrison Formation: Sedimentary Geology, 167: 297–308
Véase también
| Control de autoridades |
|
|---|
 Datos: Q57612918
Datos: Q57612918 Multimedia: Maraapunisaurus / Q57612918
Multimedia: Maraapunisaurus / Q57612918