Carcharocles megalodon
El megalodón o megalodonte (Carcharocles megalodon; del gr. μέγας megas ‘grande’, raíz megal-, y ὀδούς odoús ‘diente’, raíz odont-) es una especie extinta de tiburón que vivió entre hace 19,8 y 2,6 millones de años, aproximadamente, durante el Cenozoico (de inicios del Mioceno[2] hasta el final del Plioceno).[3] Anteriormente se pensaba que era miembro de la familia Lamnidae y pariente cercano del gran tiburón blanco (Carcharodon carcharias). Sin embargo, actualmente existe un consenso casi unánime de que pertenece a la familia extinta Otodontidae, que se separó de la ascendencia del gran tiburón blanco durante el Cretácico temprano.
| Megalodón | ||
|---|---|---|
| Rango temporal: 19,8 Ma - 2,6 Ma Burdigaliense (Mioceno) - Piacenziense (Plioceno) | ||
 Modelo de las mandíbulas del megalodón en el Museo Americano de Historia Natural. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Subfilo: | Vertebrata | |
| Clase: | Chondrichthyes | |
| Subclase: | Elasmobranchii | |
| Superorden: | Selachimorpha | |
| Orden: | Lamniformes | |
| Familia: | †Otodontidae | |
| Género: | †Carcharocles | |
| Especie: |
Carcharocles megalodon u Otodus megalodon (Agassiz, 1843)[1] | |
| Sinonimia | ||
Lista de sinónimos Género Carcharias
Género Carcharocles
Género Carcharodon
Género Megaselachus
Género Procarcharodon
Género Otodus
Género Selache
| ||

Si bien C. megalodon es considerado como uno de los mayores y más poderosos depredadores en la historia de los vertebrados,[4] los científicos difieren sobre si se habría parecido más a una versión más robusta del gran tiburón blanco, el tiburón peregrino (Cetorhinus maximus) o el tiburón tigre de arena (Carcharias taurus). La mayoría de las estimaciones del tamaño del megalodonte se extrapolan de los dientes; con estimaciones de longitud máxima de hasta 18 metros y estimaciones de longitud promedio de 10,5 metros. Las estimaciones sugieren que sus grandes mandíbulas podrían ejercer una fuerza de mordida de hasta 110,000 a 180,000 newtons. Los restos fósiles indican que este tiburón gigante tuvo una distribución cosmopolita, con áreas de cría en zonas costeras cálidas.[5] C. megalodon probablemente tuvo una influencia muy importante en la estructura de las comunidades marinas de su época.[6]
El animal se enfrentó a la competencia de los cetáceos comedores de ballenas, como Livyatan y otros cachalotes macroraptoriales y posiblemente orcas ancestrales más pequeñas. Como el tiburón prefería las aguas más cálidas, se cree que el enfriamiento oceánico asociado con el inicio de las edades de hielo, junto con la disminución de los niveles del mar y la pérdida resultante de áreas de cría adecuadas, también pudieron contribuir a su disminución. Una reducción en la diversidad de las ballenas barbadas y un cambio en su distribución hacia las regiones polares pudo reducir la fuente primaria de alimentos del megalodonte. Un estudio de 2019 revisó evidencia más reciente que sugiere que la competencia del gran tiburón blanco moderno también pudo contribuir a la extinción del megalodonte, junto con la fragmentación del rango que resulta en una extinción gradual y asíncrona como resultado del enfriamiento de los océanos hace unos 3.6-4 millones de años, mucho antes de lo que se suponía anteriormente. La extinción del tiburón parece haber afectado a otros animales; por ejemplo, el tamaño de las ballenas barbadas aumentó significativamente después de que el tiburón desapareciese.
Descubrimiento
Las glossopetræ
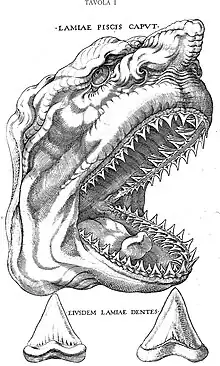
De acuerdo a las ideas del Renacimiento, los gigantescos dientes triangulares fósiles que frecuentemente se hallaban inmersos en formaciones rocosas, se consideraron lenguas petrificadas (en latín glossopetræ) de dragones y serpientes. Esta interpretación fue corregida en 1667 por un naturalista danés, Nicolás Steno, quien las reconoció como pertenecientes a tiburones antiguos, basándose en la disección que realizó de una cabeza de tiburón actual.[7] Steno describió sus hallazgos en un estudio, Disección de la cabeza de un tiburón, que contenía una ilustración de la cabeza del tiburón actual junto a dos glossopetrae, para mostrar y destacar su similitud con los dientes de este animal.[8] Según John Maisey, la lámina de Steno corresponde a la primera ilustración conocida de fósiles de C. megalodon.[9] Por otro lado, este estudio constituye la primera interpretación certera sobre el origen orgánico de los fósiles.[10]
Identificación
El naturalista suizo Louis Agassiz le dio a este tiburón su nombre científico, Carcharodon megalodon, en 1835,[11] en su trabajo de investigación Recherches sur les poissons fossiles[12] (investigaciones sobre los peces fósiles), que completó en 1843. Como los dientes de C. megalodon son morfológicamente similares a los del gran tiburón blanco, Agassiz asignó la especie al género Carcharodon.[11] Popularmente es apodado tiburón megadiente,[13] tiburón blanco gigante[14] o incluso tiburón monstruo.[15]
Hubo una aparente descripción de este tiburón en 1881, en que se lo clasificó como Selache manzonii.[16]
Fósiles
Carcharodon megalodon está representado en el registro fósil principalmente por dientes y centros de vértebras.[13] Como en los demás tiburones, el esqueleto de Carcharodon. megalodon estaba formado de cartílago más que de hueso; esto resulta en una pobre preservación de los especímenes encontrados.[17] Sin embargo, los restos fósiles de C. megalodon indican que tenía los centros vertebrales densamente calcificados.[18]
Dientes fósiles


Los fósiles más comunes de C. megalodon son sus dientes. Las características diagnósticas de los dientes de C. megalodon incluyen: forma triangular,[5] estructura robusta,[13] gran tamaño,[5] un borde finamente aserrado,[5] y el cuello (base de la corona) con una visible forma de letra v.[5] Los dientes de C. megalodon miden 18,5 centímetros en altura perpendicular y 13,5 centímetros en longitud diagonal, y son los mayores en tamaño de cualquier especie conocida de tiburón.[19]
Vértebras
Se han encontrado algunas vértebras fósiles de C. megalodon.[9] El ejemplo más notable es una columna vertebral parcialmente preservada de un espécimen de C. megalodon, la cual fue desenterrada en la Cuenca de Amberes, Bélgica por M. Leriche en 1926. Este espécimen comprendía varios centros vertebrales, midiendo el mayor unos 155 milímetros de diámetro.[13] Sin embargo, algunos paleontólogos han afirmado que podrían esperarse centros vertebrales considerablemente mayores de C. megalodon.[13] Bendix-Almgreen (1983) publicó el hallazgo de veinte centros vertebrales sin conexión anatómica, pero muy próximos entre sí, procedentes de las arcillas de la Formación Gram (Dinamarca). Estos restos, de 100 a 230 milímetros de diámetro, mostraban una densa calcificación y estructura concéntrica.[18]
Distribución paleogeográfica y edad
Se conocen fósiles de C. megalodon procedentes de diferentes partes del mundo, que incluyen a Europa,[9] América del Norte,[13] América del Sur,[9][13] Puerto Rico,[20] Cuba,[21] Jamaica,[22] Australia,[23] Nueva Zelanda,[19] Japón,[9][13] África,[9][13] Malta,[19] Granadinas,[24] Islas Canarias,[25] e India.[9] Algunos dientes de C. megalodon se descubrieron en regiones muy alejadas de los continentes (por ejemplo, en la Fosa de las Marianas en el Océano Pacífico).[19]
Aunque los restos más antiguos de C. megalodon fueron reportados de estratos del Oligoceno tardío, hace cerca de 28 millones de años.[26][27] una fecha más aceptada para el origen de la especie es durante el Mioceno Medio, hace unos 15.9 millones de años.[3] A pesar de que el registro fósil de C. megalodon por lo general está ausente en estratos posteriores al límite superior del Terciario,[13] existe registro más reciente en estratos del Pleistoceno.[28] Se cree que C. megalodon se extinguió en el final del Plioceno, probablemente hace unos 2.6 millones de años;[3] se piensa por lo tanto que los dientes post-pliocénicos reportados de C. megalodon son en realidad fósiles reelaborados.[17]
Anatomía
Entre las especies existentes, se considera que el gran tiburón blanco es la más análoga a C. megalodon.[5] La carencia de esqueletos fósiles bien preservados de C. megalodon ha forzado a los científicos a basarse en la morfología del gran tiburón blanco para inferir su aspecto y estimar su tamaño.[13]
Estimación de tamaño
Debido a lo fragmentario de los restos hallados hasta ahora, estimar el tamaño de C. megalodon se enfrenta a una incertidumbre elevada.[19] Sin embargo, la comunidad científica reconoce que C. megalodon superaba en tamaño al tiburón ballena (Rhincodon typus). Los investigadores han dirigido sus estudios hacia dos aspectos del tamaño: la longitud total (LT) y la masa corporal (MC).
Estimaciones de longitud total
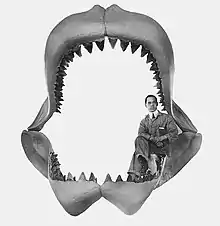
El primer intento de reconstruir la mandíbula de este tiburón fue realizado por el profesor Bashford Dean en 1909. De las dimensiones de la mandíbula reconstruida, se supuso que C. megalodon podría haberse aproximado a los 30 metros de longitud total (LT) pero, a la luz de nuevos descubrimientos fósiles y avances científicos sobre la comprensión de los vertebrados, se considera ahora que esta reconstrucción fue inexacta,[29] por dos razones principales: (1) el relativamente pobre conocimiento de la dentición de C. megalodon en la época de Dean, y (2) estructuras musculares inadecuadas.[29] Los expertos sugieren que una versión rectificada del modelo de mandíbula de C. megalodon de Bashford Dean podría tener cerca del 70 % de su tamaño original, un tamaño más coherente con los hallazgos modernos.[29] Para corregir los errores, los científicos, ayudados por nuevos hallazgos fósiles de C. megalodon y un conocimiento mejorado de la anatomía de sus más cercanos análogos modernos, introdujeron más métodos cuantitativos para estimar el tamaño basándose en las relaciones estadísticas entre el tamaño de los dientes y la longitud corporal en el gran tiburón blanco.[13][29] Algunos de los métodos son mencionados a continuación.
Método propuesto por John E. Randall
En 1973, un ictiólogo de Hawaii, John E. Randall, presentó un método para estimar la longitud total del gran tiburón blanco.[30] El método propuesto es representado en forma de un gráfico trazado, el cual demuestra la relación entre la altura de la corona (la distancia vertical de la hoja del diente desde la base de la parte de esmalte del diente hasta su punta) del mayor de los dientes de la mandíbula superior del gran tiburón blanco y su longitud total.[29][30] Randall también extrapoló este método a la longitud total de C. megalodon.[30] Randall citó dos dientes de C. megalodon en su trabajo: (1) el espécimen número 10356 del Museo Americano de Historia Natural y (2) el espécimen número 25730 de Museo Nacional de Estados Unidos, los cuales tenían una altura de la corona de 115 mm. y 117.5 mm. respectivamente.[30] Estos dientes produjeron una longitud total de cerca de 13 metros.[29][30] En 1991, dos expertos en tiburones, Richard Ellis y John E. McCosker, señalaron un defecto en el método de Randall.[13] De acuerdo a ellos, la altura de la corona dental de los tiburones no necesariamente se incrementa en proporción a la longitud total del animal. Esta observación llevó a nuevas propuestas para dar con métodos más exactos para determinar el tamaño del gran tiburón blanco y especies similares.[13]
Método propuesto por Gottfried et al.
En 1996 tres científicos —Michael D. Gottfried, Leonard J. V. Compagno y S. Curtis Bowman— después de haber realizado una minuciosa investigación y escrutinio de 73 especímenes del gran tiburón blanco, propusieron una relación linear conservadora entre la máxima altura del mayor de los dientes superiores (DS) y la longitud total en dicho tiburón.[13][27] La relación propuesta es: la longitud total (LT) en metros = − (0.22) + (0.096) × [máxima altura de DS (milímetros)].[13][27] Gottfried y colegas (1996) también extrapolaron este método para estimar la longitud total de C. megalodon. El mayor diente de C. megalodon al alcance de este equipo era un espécimen de segundo diente anterior superior, cuya máxima altura era de 168 mm. Este diente había sido descubierto por Compagno en 1993, y produjo una longitud total estimada en 15.9 metros.[13] Sin embargo, rumores de dientes mayores de C. megalodon persistían en esa época.[13] La máxima altura dental para este método se mide como una línea vertical desde la punta de la corona dental a la parte baja de los lóbulos de la raíz, paralelo al largo eje del diente.[13] Dicho de otra forma, la máxima altura del diente es la altura perpendicular.[31]
Método propuesto por Clifford Jeremiah
En 2002, el investigador de tiburones Clifford Jeremiah propuso un método para estimar la longitud total del gran tiburón blanco y especies parecidas incluyendo a C. megalodon.[19] El investigador David Ward ha asegurado que este método está basado en un sólido principio que funciona bien con muchos tiburones grandes.[19] El método en sí propone: «por cada centímetro de anchura de la raíz de un diente superior anterior, hay aproximadamente 4.5 pies (1.3 metros) de tiburón». Jeremiah señaló que el perímetro de la mandíbula de un tiburón es directamente proporcional a su longitud total, siendo la anchura de las raíces de los dientes mayores una aproximación para estimar el perímetro de la mandíbula.[19] El mayor diente en posesión de Jeremiah tenía una raíz de cerca de 12 cm, lo cual indica una longitud total de 15.5 metros.[19]
Método propuesto por Kenshu Shimada
En 2002, el paleontólogo Kenshu Shimada de la Universidad DePaul propuso una relación linear entre la altura de la corona (AC) y la longitud total (LT) en los grandes tiburones blancos después de realizar análisis anatómicos de varios ejemplares.[32] En términos estadísticos, esta relación se expresa como: la longitud total (LT) en cm. = a + bx, donde a es una constante, b es la pendiente de la línea, y x es la altura de la corona (AC) del diente en milímetros. Esta relación permite predecir la longitud total sobre la base de la altura de la corona de cualquier diente en el gran tiburón blanco.[5][32] Para este método, la altura de la corona del diente es medida como la máxima altura vertical del esmalte en el lado labial.[32] Shimada señaló que los métodos antes propuestos estaban basados en una débil evaluación de la homología dental.[32] Shimada indicó que la tasa de crecimiento entre la corona y la raíz no es isométrica, y este factor debe ser tomado en cuenta en su modelo.[32] Más aún, esta relación propuesta puede también ser usada para predecir la longitud total de tiburones que sean morfológicamente similares a gran tiburón blanco (como C. megalodon).[5][32] Usando este modelo, el diente superior anterior (con una altura máxima de 168 mm.) citado por Gottfried y colaboradores (1996) corresponde a una longitud total de 15.1 metros.[32]
En 2010, varios investigadores de tiburones, como Catalina Pimiento, Dana J. Ehret, Bruce J. MacFadden y Gordon Hubbell, estimaron la longitud total de C. megalodon sobre la base del método de Shimada.[5] Entre los especímenes hallados en la formación Gatún de Panamá, el ejemplar número 237956 produjo una longitud total de 16.8 metros.[5]
Consenso


En la década de 1990, algunos biólogos marinos (como Patrick J. Schembri y Staphon Papson) opinaron que C. megalodon pudo alcanzar un máximo de entre 24 a 25 metros de longitud total.[33][34] La anterior estimación de la longitud total de C. megalodon quizás fuera sea tan descabellada. Sin embargo, Gottfried y sus colaboradores (1996) propusieron que C. megalodon pudo medir como mucho 20,3 metros de longitud.[4][13][35] Actualmente el consenso es que este tiburón gigante medía más de 16 metros.[5][4][19][35]
Mayores especímenes conocidos
Gordon Hubbell de Gainesville en Estados Unidos, dispone de un diente superior anterior de C. megalodon cuya altura máxima es 184,15 mm.[19] Adicionalmente, una reconstrucción de la mandíbula de C. megalodon contiene un diente cuya máxima altura reportada es 195,833 mm.[36] Esta mandíbula reconstruida fue realizada por el difunto cazador de fósiles Vito Bertucci,[36] quien era conocido como el "Hombre Megalodon".[19][36]
Estimaciones de masa corporal
Gottfried y colaboradores (1996) también presentaron un método para determinar la masa corporal del gran tiburón blanco después de estudiar los datos de la relación de longitud-masa corporal de 175 especímenes en varias etapas de crecimiento y lo extrapolaron para estimar la masa corporal de C. megalodon. El método propuesto es: masa en kilogramos = 3,29E−06[LT en (metros)3.174].[13] De acuerdo con este modelo, un C. megalodon de 15,9 metros pudo haber tenido una masa corporal de cerca de 47 toneladas,[4][13] un C. megalodon de 17 metros sería de cerca de 59 toneladas,[13] y un C. megalodon de 20,3 metros tendría una masa de 103 toneladas.[4][13] Consecuentemente, C. megalodon es considerado como el mayor tiburón que haya vivido,[5] y está entre los mayores peces conocidos que hayan existido.[13]
Dentición y mecánica de la mandíbula

Un equipo de científicos japoneses, T. Uyeno, O. Sakamoto y H. Sekine, descubrieron y excavaron restos parciales de C. megalodon, con una colección de dientes casi completa hallada en asociación, en Saitama, Japón en 1989.[9] Otra dentición completa asociada de C. megalodon fue excavada en la formación Yorktown de Lee Creek, Carolina del Norte en Estados Unidos y sirvió como la base de una reconstrucción de la mandíbula de C. megalodon del Museo Americano de Historia Natural en Nueva York.[13] Estos hallazgos de dientes asociados resolvieron la incertidumbre de cuántos dientes podrían estar en cada hilera de las mandíbulas de C. megalodon, posibilitando reconstrucciones más exactas de las mismas. Más denticiones asociadas de C. megalodon también se han hallado en los últimos años. Basándose en estos descubrimientos, dos científicos, S. Applegate y L. Espinosa, publicaron una fórmula dental teórica (una representación de la dentición de un animal con respecto a los tipos de dientes y su disposición dentro de la mandíbula del animal) para C. megalodon en 1996.[9][13] Las reconstrucciones modernas de la mandíbula de C. megalodon están basadas en dicha fórmula dental: [9]

Como se manifiesta en la fórmula dental, C. megalodon tenía cuatro clases de dientes en sus mandíbulas.[9]
- Anterior - A
- Intermedio - I (En el caso de C. megalodon, este diente parece ser técnicamente un superior anterior y es denominado como "A3" debido a que es bastante simétrico y no apunta medialmente (el lado del diente se inclina hacia la línea media de las mandíbulas donde las mandíbulas izquierda y derecha se encuentran), pero este diente aun así está diseñado como un diente intermedio.[11] Sin embargo, en el caso del gran tiburón blanco, el diente intermedio sí apunta medialmente. Este aspecto ha sido frecuentemente puesto de relieve en el debate Carcharodon versus Carcharocles sobre el megalodon y favorece a los proponentes de su clasificación en Carcharocles.)
- Lateral - L
- Posterior - P

C. megalodon tenía una dentición muy robusta,[13] y tenía un total de cerca de 276 dientes en sus mandíbulas, que abarcan 5 hileras. (Véase "enlaces externos" más adelante en el texto)
Los paleontólogos sugieren que un C. megalodon muy grande tendría mandíbulas de cerca de 2 metros de un lado a otro.[19]
Fuerza de mordida
En 2008, un equipo de científicos dirigido por Stephen Wroe llevó a cabo un experimento para determinar la fuerza de mordida de C. megalodon; los resultados indican que tenía una de las más poderosas fuerzas de mordida de la historia.[4] Un C. megalodon de 15,9 metros de largo era capaz de ejercer una fuerza de mordida estimada en 108.514 newton,[4] y un C. megalodon de 20,3 metros de largo sería capaz de ejercer una mordida estimada en 182.201 newton.[4]
La fuerza de mordida de C. megalodon, en su tamaño máximo estimado, es cerca de 28 veces mayor que la del pez Dunkleosteus en 5,3 kilonewton,[4][37] cerca de 10 veces mayor que la del gran tiburón blanco en 18 kilonewton,[4] cerca de 5 veces mayor que la del Tyrannosaurus rex en 31 kilonewton,[4] y también es mayor que la del pliosaurio Pliosaurus funkei en 150 kilonewton.
Adicionalmente, Wroe y colegas (2008) señalaron que los tiburones también se sacuden de lado a lado cuando se alimentan, amplificando las fuerzas postcraneales generadas. Por lo tanto las fuerzas totales experimentadas por las presas son probablemente más altas que las fuerzas estimadas a través del experimento.[4] La extraordinaria fuerza de mordida de C. megalodon debe ser considerada en el contexto del gran tamaño de este depredador extinto y a que la evidencia paleontológica sugiere que C. megalodon era un activo depredador de grandes ballenas.[4]
Parámetros funcionales de los dientes
Los dientes excepcionalmente robustos de C. megalodon son aserrados,[11][19] lo cual pudo haber mejorado su eficiencia al cortar la carne de sus presas. El paleontólogo Dr. Bretton K. Kent de la Universidad de Maryland sugiere que estos dientes son muy gruesos para su tamaño, con coeficientes mucho menores de esbeltez y de resistencia a la flexión. Tenían también raíces sustancialmente mayores comparadas a la altura total del diente, dándoles una gran ventaja mecánica. Dientes con estas características no sólo son buenas herramientas de corte, sino también son muy adecuados para sujetar a presas poderosas y rara vez se romperían - incluso cortando a través de los huesos.[38]
Anatomía del esqueleto

Aparte de estimar el tamaño de C. megalodon, Gottfried y colegas (1996) también trataron de determinar la esquemática del esqueleto entero de C. megalodon.[13]
Estructura de la mandíbula
Para tener apoyo funcional para su enorme y robusta dentición, las mandíbulas de C. megalodon debieron de haber sido enormes, gruesas y más fuertemente desarrolladas que las del gran tiburón blanco, el cual posee una dentición más grácil en comparación.[13] Las mandíbulas fuertemente desarrolladas debieron darle cierta apariencia de ojos hundidos.[13]
Condrocráneo
El condrocráneo de C. megalodon debió de tener una apariencia más compacta y robusta que el del gran tiburón blanco, para así reflejar funcionalmente sus mandíbulas y dentadura más macizas en comparación.[13]
Aletas
Las aletas de C. megalodon debieron haber sido muy probablemente proporcionalmente más grandes y gruesas comparadas con las del gran tiburón blanco debido a que las aletas relativamente grandes son necesarias para la propulsión y control de movimientos en un tiburón mayor.[13]
Esqueleto axial
A través del escrutinio de las vértebras parcialmente preservadas del espécimen de C. megalodon hallado en Bélgica, parece ser que C. megalodon tenía un número mayor de vértebras que los hallados en cualquier espécimen grande de los tiburones conocidos.[13] Sólo el número de vértebras del gran tiburón blanco se aproxima en cantidad, simbolizando los cercanos vínculos anatómicos entre las dos especies.[13]
Reconstrucción del esqueleto completo
Sobre la base de las características mencionadas antes, Gottfried y colaboradores (1996) reconstruyeron un esqueleto completo de un individuo juvenil de C. megalodon de 11,5 metros de longitud, el cual se exhibe en el Museo Marino Calvert en la isla Solomons (Maryland, Estados Unidos).[13][39][40][13] El equipo hizo hincapié en que las diferentes proporciones al compararlo con el esqueleto de un gran tiburón blanco, están basadas en los mismos patrones ontogenéticos de crecimiento del propio gran tiburón blanco actual.[13]
Consideraciones paleoecológicas
Distribución y hábitat
Los tiburones, especialmente las especies grandes, son organismos altamente móviles con un complejo ciclo vital y amplia distribución geográfica.[5] Los registros fósiles de C. megalodon indican que eran cosmopolitas,[19] y comúnmente aparecen en latitudes subtropicales a templadas.[13] Antes de la formación del Istmo de Panamá, los mares eran relativamente cálidos.[41] Esto hizo posible para esta especie vivir en todos los océanos del mundo.
C. megalodon tenía la suficiente flexibilidad de comportamiento para habitar un amplio rango de ecosistemas marinos (aguas costeras poco profundas,[42] afloramientos costeros,[42] lagunas costeras pantanosas,[42] litorales arenosos,[42] y ambientes de aguas profundas de alta mar[19]), exhibiendo un modo de vida transitorio.[42] Los adultos de C. megalodon no eran abundantes en ambientes de aguas costeras poco profundas,[42] y mayormente acechaban mar adentro. C. megalodon pudo haberse movido entre aguas costeras y oceánicas, particularmente en distintas etapas de su vida.
Interacción con las presas

Los tiburones son generalmente depredadores oportunistas. Sin embargo, los científicos proponen que C. megalodon era "sin duda el más formidable carnívoro que haya existido".[4] Su gran tamaño,[4] capacidad de nado rápido,[40] y poderosas mandíbulas se conjuntaban en un formidable aparato predatorio,[4][13] haciéndolo un superdepredador con la capacidad de consumir un amplio espectro de presas.
La evidencia fósil indica que C. megalodon cazaba cetáceos como delfines,[13] pequeñas ballenas,[9] (incluyendo cetotéridos[17] tales como Piscobalaena,[43] escualodóntidos,[42] y Odobenocetops[44]), así como grandes ballenas,[45] (incluyendo a cachalotes,[19][46] ballenas de Groenlandia,[47] y rorcuales[45][48]), pinípedos,[17][24][49] marsopas,[19] sirenios,[42][50] y grandes tortugas marinas.[42]
Los mamíferos marinos eran blancos regulares de C. megalodon. Muchos huesos de ballenas se han encontrado con grandes marcas de mordidas (cortes profundos) hechos por dientes que encajan con los de C. megalodon,[9][13] y varias excavaciones han revelado que los dientes de C. megalodon yacen cerca de restos masticados de ballenas,[13][39] y a veces en directa asociación con ellos.[14] También hay evidencia fósil de interacciones entre C. megalodon y pinípedos.[24] En una interesante observación, un diente de C. megalodon de 127 mm. fue hallado muy cerca de un hueso del oído mordido de un león marino.[49]

C. megalodon se enfrentó a un ambiente muy competitivo durante su existencia.[6] Sin embargo, C. megalodon, estando en lo alto de la cadena alimenticia,[51] probablemente tuvo un profundo impacto en las comunidades marinas.[13][6][52] La evidencia fósil indica una correlación entre el surgimiento de C. megalodon y la extensa diversificación de los cetáceos por el mundo.[13][6] Los C. megalodon jóvenes preferían regiones donde los cetáceos pequeños eran abundantes, y los adultos preferían regiones donde abundaran especies mayores.[13] Dichas preferencias pueden haberse desarrollado poco después de su aparición en el Oligoceno.[13] Adicionalmente, C. megalodon fue contemporáneo de odontocetos macrodepredadores (particularmente cachalotes carnívoros y escualodóntidos), los cuales probablemente estaban entre los principales depredadores marinos de su tiempo,[15][6] y eran una competencia.[15][53] En respuesta a la competencia de los tiburones gigantes superdepredadores, los odontocetos carnívoros pudieron haber desarrollado algunas adaptaciones defensivas; algunas especies se volvieron cazadores en manada,[15][54] mientras algunas especies alcanzaron tamaños gigantescos, como Livyatan melvillei.[6][55] Aun así, marcas de mordida en restos fósiles de odontocetos indican que fueron presa de los tiburones gigantes.[42][46] Además, la evidencia fósil indica que C. megalodon desarrolló la capacidad de atacar ballenas grandes.[4][13] A finales del Mioceno, los cachalotes superdepredadores desaparecieron del registro fósil y dejaron un vacío ecológico.[6]
Como otros tiburones, C. megalodon también pudo haber sido piscívoro.[17][40] La evidencia fósil indica que otras especies notables de tiburones superdepredadores (como el gran tiburón blanco) respondieron a la presión de la competencia de C. megalodon evitando las zonas que este habitaba.[13] C. megalodon probablemente tenía también tendencia al canibalismo.[56] El consumo de presas por parte de C. megalodon parece haber variado según la edad y el lugar, como ocurre con el gran tiburón blanco actual. Es posible que la población adulta de C. megalodon que habitaba a lo largo de las costas de Perú prefiriera cazar con frecuencia a las ballenas cetotéridas de 2.5 - 7 metros de longitud y a otros animales de menor talla que ellos mismos, en lugar de las ballenas con el mismo rango de tamaño que los megalodontes.[43] Entretanto, los juveniles probablemente tendrían una dieta que consistiría sobre todo de peces.[57]
Estrategias de alimentación
Los tiburones frecuentemente emplean complejas estrategias de caza para a abordar a grandes presas. Algunos paleontólogos sugieren que las estrategias de caza del gran tiburón blanco pueden ofrecer pistas de como el C. megalodon puede haber cazado presas inusualmente grandes (como ballenas).[15] Sin embargo, la evidencia fósil sugiere que C. megalodon empleaba estrategias de caza más efectivas contra grandes presas comparadas por las usadas por el gran tiburón blanco.[38]
Los paleontólogos han realizado un estudio de los fósiles para determinar los patrones de ataque de C. megalodon en sus presas.[38] Un espécimen en particular del Mioceno — los restos de una ballena de 9 metros de largo (de un taxón sin identificar) — proporcionó la primera oportunidad de analizar cuantitativamente el comportamiento predatorio de C. megalodon.[38] Este cazador se enfocaba principalmente en atacar las partes con más hueso (por ejemplo, hombros, aletas, costillas y la columna vertebral) de la presa,[38] las cuales los grandes tiburones blancos generalmente evitan.[38] Bretton Kent indica que C. megalodon trataba de destrozar los huesos y dañar los delicados órganos (el corazón y los pulmones) alojados en la caja torácica de las presas.[38] Con un ataque pudo haber inmovilizado a la presa, la cual moriría rápidamente debido a las heridas en estos órganos vitales.[38] Estos hallazgos también hacen claro por qué este tiburón prehistórico necesitaba unos dientes más robustos que los de los grandes tiburones blancos.[38] Además, los patrones de ataque podrían diferir según el tamaño de las presas.[15] Los restos fósiles de algunos cetáceos pequeños (como los cetotéridos) sugieren que eran embestidos con gran fuerza desde abajo antes de ser muertos y devorados.[15]
Durante el Plioceno aparecieron cetáceos más grandes y avanzados.[58] C. megalodon aparentemente refinó sus estrategias de caza para lidiar con estas grandes ballenas. Numerosos huesos de aletas fosilizados (segmentos de las aletas pectorales) y de las vértebras caudales de grandes ballenas del Plioceno se han hallado con marcas que fueron causadas por ataques de C. megalodon.[4] Esta evidencia paleontológica sugiere que C. megalodon podría tratar de inmovilizar una ballena grande destrozando sus estructuras de propulsión antes de matarla y alimentarse de ella.[4]
Áreas de cría

La evidencia fósil sugiere que las áreas de cría preferidas por C. megalodon eran ambientes de aguas costeras cálidas, donde los peligros potenciales eran menores y las fuentes de alimento abundantes.[5] Se han identificado algunos sitios de cría por fósiles encontrados en la Formación Gatún de Panamá,[5] la Formación Calvert en Maryland,[5] el Banco de Concepción en las Islas Canarias[59] y la Formación Bone Valley de Florida.[5] Como en el caso de muchos tiburones, C. megalodon probablemente también daba a luz crías vivas. El tamaño de los dientes de C. megalodon neonatos indica que las crías medían entre 2 a 3 metros de largo al nacer.[5][19] Sus preferencias dietarias exhibían un cambio ontogenético.[13] Los jóvenes C. megalodon comúnmente cazaban peces,[5] tortugas marinas,[42] dugones,[19] y pequeños cetáceos;[13] los C. megalodon maduros se desplazaban mar adentro en áreas frecuentadas por grandes cetáceos, que eran su fuente principal de alimento. Sin embargo, hay un caso excepcional de un supuesto C. megalodon joven que trató de atacar a una ballena grande, un rorcual. Este incidente ocurrió en el Plioceno, hace 3-4 millones de años, en Carolina del Norte, EE. UU.[60]
Extinción
La causa o causas de la extinción de C. megalodon están aún bajo investigación,[15] sin embargo hace cerca de 3,1 millones de años ocurrió un importante evento geológico — el cierre del paso marítimo de Centroamérica,[61] que causó significativos cambios ambientales y en las faunas alrededor del mundo,[61] incluyendo probablemente la extinción de C. megalodon.[9][61]
Enfriamiento oceánico y caída en los niveles del mar
El cierre del Istmo de Panamá cambió fundamentalmente la circulación oceánica global.[9][62] Este evento permitió el inicio a una glaciación en el Hemisferio Norte,[62] y más tarde, también facilitó el enfriamiento del planeta entero.[62] Adicionalmente, la expansión de la glaciación durante el Plioceno retuvo grandes volúmenes de agua en forma de capas de hielo, lo que resultó en una significativa caída en los niveles del mar.[41] A consecuencia de esto, durante el Plioceno tardío y el Pleistoceno hubo varias edades del hielo,[63][64] que enfriaron los océanos significativamente.[9] La principal razón citada para la extinción del C. megalodon es el declive en las temperaturas de los océanos a escala global durante el Plioceno.[9][39][65] Esta tendencia al enfriamiento impactó de forma adversa a C. megalodon, ya que prefería aguas cálidas,[13][65] y como resultado se volvió una especie extremadamente rara hasta su extinción final durante el Pleistoceno.[65] No hay evidencia fósil de C. megalodon en regiones alrededor del mundo en las que la temperatura del agua había caído significativamente durante el Plioceno.[13] Más aún, estos cambios oceanográficos pueden haber limitado muchos de los sitios de aguas cálidas que servían de área de cría para C. megalodon, obstaculizando el mantenimiento de su población.[9] Las áreas de cría son esenciales para el mantenimiento de una especie.[66]
Declive de las fuentes de alimentos
Los cetáceos alcanzaron su mayor diversidad durante el Mioceno,[13] con cerca de 20 géneros reconocidos en comparación a los seis géneros vivientes.[67] Dicha diversidad representaba el marco ideal para mantener a un superdepredador como C. megalodon.[13] Después del cierre del Istmo de Panamá, muchas especies de cetáceos se extinguieron,[61][68] y muchas especies sobrevivientes desaparecieron de los trópicos.[69] Los patrones de migración de las ballenas del Plioceno se han reconstruido a partir del registro fósil, sugiriendo que muchas especies sobrevivientes mostraron una tendencia a dirigirse hacia las regiones polares.[41] Las temperaturas del agua más fría durante el Plioceno impidieron a C. megalodon llegar a las regiones polares, y las mayores presas ya no estaban "dentro del rango" de C. megalodon después de dichas migraciones.[9][13][17][69] Estos sucesos disminuyeron la fuentes de alimentos de C. megalodon en las regiones que éste habitaba en el Plioceno, principalmente el latitudes medias a bajas.[9][45][69] C. megalodon estaba adaptado a un estilo de vida especializado,[39] el cual fue perturbado como resultado de estos cambios.[39] La extinción de C. megalodon se correlaciona con el declive de muchos linajes de misticetos pequeños; es posible que fuera muy dependiente de ellos como fuente de alimento.[43] Además, la escasez de presas en los trópicos durante los tiempos del Plio-Pleistoceno puede haber impulsado el canibalismo en los C. megalodon.[15] Los individuos jóvenes estarían en un creciente riesgo de ataques de los adultos durante las épocas de hambruna.[15]
Competencia ecológica con nuevas especies
La aparición y amplia distribución de los grandes delfínidos depredadores del género Orcinus durante el Plioceno,[70] llenó probablemente el vacío ecológico dejado a fines del Mioceno por la desaparición de los cachalotes superdepredadores.[6] Una interpretación minoritaria indica que estos delfínidos pudieron haber superado la competencia ecológica con los últimos representantes de C. megalodon en el nicho de depredador superior.[71] Sin embargo, muchos expertos han inferido que factores como la tendencia al enfriamiento en los océanos y la disminución de presas durante el Plioceno pueden haber jugado un papel muy significativo en el declive y en la extinción de C. megalodon.[15]
El registro fósil indica que estos delfínidos aparecen comúnmente en latitudes altas durante el Plioceno,[70] indicando que pudieron hacer frente a las condiciones de aguas frías que se estaban volviendo predominantes, aunque también se han encontrado en los trópicos (como Orcinus sp. en Sudáfrica).[70] Grandes marcas de mordidas en restos fósiles de delfínidos indican que C. megalodon los cazaba.[13] Sin embargo, el paleontólogo Albert Sanders sugiere que C. megalodon se había vuelto demasiado grande como para sustentarse de las fuentes de alimentación disponibles en los trópicos.[66]
Múltiples causas
El consenso de los expertos ha señalado que factores como el enfriamiento de los océanos y la disminución de las fuentes de comida durante el Plio-Pleistoceno constituyeron un factor significativo en el declive de la especie.[15]
Un estudio publicado en 2016 de la distribución y el ambiente del megalodonte a través del tiempo de su existencia indicó que los factores bióticos, es decir la competencia con nuevos depredadores combinada con la reducción del número de sus presas, fueron las causas principales de su extinción.[2] Un análisis de la distribución y abundancia de este tiburón a lo largo del Mioceno y el Plioceno no encontró correlación con los patrones de calentamiento y enfriamiento del océano. Su declive global comenzó en el punto en que había alcanzado su máximo rango geográfico. Se halló que C. megalodon habitaba océanos con una temperatura de 12 a 27 °C, indicando que la extensión global de hábitats apropiados no habría sido muy afectada por los cambios de temperatura que estaban ocurriendo. Esto puede reflejar además que C. megalodon, como otros grandes tiburones, era parcialmente endotérmico.[2] Sin evidencia de que las restricciones de temperatura fueran importantes, los factores bióticos que involucraron la disminución de la diversidad de cetáceos, en especial de ballenas barbadas, y el incremento de la competencia con los grandes tiburones blancos, los fiseteroideos y delfínidos superdepredadores parecen haber sido los principales responsables del declive y desaparición del megalodonte.[2][72]
El tamaño corporal de las ballenas con barbas se incrementó significativamente tras la extinción del megalodonte[3] (aunque esto podría deberse a causas más relacionadas con el clima)[73] Otros superdepredadores parecen haberse beneficiado con la desaparición de esta formidable especie.[65]
Taxonomía
Incluso después de décadas de investigación y escrutinio, la controversia sobre la filogenia de C. megalodon aún persiste.[11][74] Varios investigadores de tiburones (como J. E. Randall, A. P. Klimley, D. G. Ainley, M. D. Gottfried, L. J. V. Compagno, S. C. Bowman y R. W. Purdy) insisten que C. megalodon es un pariente cercano del gran tiburón blanco. Sin embargo, varios otros expertos (como D. S. Jordan, H. Hannibal, E. Casier, C. DeMuizon, T. J. DeVries, D. Ward y H. Cappetta) rechazan esa propuesta y citan la evolución convergente como la razón principal de la similitud en los dientes de ambos. Los argumentos de los que apoyan al género Carcharocles para C. megalodon parecen haber ganado un notable apoyo.[39] Sin embargo, la asignación taxonómica original aún tiene amplia aceptación.[11]
Megalodonte dentro de Carcharodon
|
|
La visión tradicional es que C. megalodon debería ser clasificado dentro del género Carcharodon junto con el gran tiburón blanco. Las razones principales citadas para esta filogenia son: (1) una gradación ontogénica, mediante la cual los dientes de C. carcharias pasan de tener un aserramiento tosco en los jóvenes a uno más fino en los adultos, siendo los últimos más parecidos a los de C. megalodon; (2) la similitud morfológica de los dientes de los C. megalodon jóvenes a los de C. carcharias; (3) un segundo diente anterior simétrico; (4) grandes dientes intermedios que se inclinan medialmente; y (5) los dientes anteriores superiores tienen la zona lingual del cuello del diente con forma de cheurón. Los partidarios de la clasificación dentro de Carcharodon para C. megalodon sugieren que C. megalodon y C. carcharias comparten un ancestro común, Palaeocarcharodon orientalis.[11][19]
Megalodonte dentro de Carcharocles
|
|
Allá por 1923, el género Carcharocles fue propuesto por dos investigadores de tiburones, D. S. Jordan y H. Hannibal, para clasificar al tiburón C. auriculatus. Más tarde, los proponentes de Carcharocles asignaron C. megalodon al género Carcharocles.[11][19] Los proponentes de Carcharocles también sugirieron que el ancestro directo de los tiburones pertenecientes al género Carcharocles, es el antiguo tiburón gigante Otodus obliquus, el cual vivió durante las épocas Paleoceno y Eoceno.[39][74] De acuerdo a los partidarios de la clasificación de C. megalodon en el género Carcharocles, la línea filogenética que da lugar a los megalodontes sería: Otodus obliquus - Otodus aksuaticus - Carcharocles auriculatus - Carcharocles angustidens - Carcharocles chubutensis - C. megalodon.[19][39] Los dientes de Carcharocles chubutensis ya muestran la pérdida de las pequeñas cúspides laterales que caracterizan los de Carcharocles angustidens, ausentes definitivamente en C. megalodon.[19][39] Incluso es posible que C. megalodon fuera una especie de Otodus. Por lo tanto, algunos investigadores lo denominan Otodus megalodon.[75]
Reconsideración del linaje megadentado de Carcharocles a Otodus
Los especialistas en tiburones han estado considerando el regreso del linaje entero de Carcharocles al género Otodus.[75] Un estudio de 1974 sobre tiburones del Paleógeno acuñó al subgénero Megaselachus, clasificando a este tiburón como Otodus (Megaselachus) megalodon, junto con O. (M.) chubutensis.[76] Una revisión de 2006 de los Chondrichthyes elevó el nombre al rango de género, reclasificando a ambas especies como Megaselachus megalodon y M. chubutensis.[77] La evidencia de que el megalodonte es un miembro de Otodus se fortaleció en 2016 con la publicación de un nuevo género relacionado, Megalolamna.[78][79] En su estudio, Shimada et al. concluyeron que reclasificar a todos los miembros de Carcharocles en Otodus es necesario para hacer a Otodus monofilético, y que Otodus es el clado hermano de Megalolamna dentro de la familia Otodontidae. A su vez dentro del género Otodus, se considera que O. obliquus es el taxón hermano del linaje de tiburones megadentados.[79]
¿El megalodón como cronoespecie?
El investigador David Ward ha elaborado aún más el proceso evolutivo de Carcharocles al implicar que este linaje, que se extiende desde el Paleoceno al Plioceno, es de un único tiburón gigante el cual cambia gradualmente a través del tiempo, sugiriendo un caso de cronoespecie.[19]
Una nueva posición evolutiva para el gran tiburón blanco
Los proponentes de Carcharocles señalaron que el gran tiburón blanco está más estrechamente relacionado al antiguo tiburón Isurus hastalis, conocido como el "mako de dientes anchos", que a C. megalodon.[11] Una razón citada por el paleontólogo Chuck Ciampaglio es que la morfometría dental (variaciones intraespecíficas de las medidas y cambios en la forma física de los dientes) de I. hastalis y C. carcharias son notablemente similares.[11] Otra razón citada es que los dientes de C. megalodon tienen sierras mucho más finas que los de C. carcharias.[11] Otra evidencia adicional, que vincula al gran tiburón blanco más cercanamente a los antiguos tiburones mako que a C. megalodon, se publicó en 2009: los fósiles de una antigua forma del gran tiburón blanco, datada en cerca de 4 millones de años, y que habían sido encontrados en el suroeste de Perú en 1988. Estos restos muestran un probable ancestro compartido por los makos modernos y los grandes tiburones blancos actuales.[74][80]
Polémicas

El paleontólogo Chuck Ciampaglio de la Universidad Estatal de Wright afirma que las similitudes entre los dientes de C. megalodon y los del gran tiburón blanco son superficiales y tienen notables diferencias morfométricas entre ambos, y que estos hallazgos son suficientes como para justificar un género aparte para C. megalodon.[11][15] Sin embargo, algunos partidarios del género Carcharodon para C. megalodon (como M. D. Gottfried y R. E. Fordyce) han provisto más argumentos para una cercana relación entre los tiburones megadentados extintos y el gran tiburón blanco.[27] Con respecto a la reciente controversia concerniente a las relaciones entre los tiburones lámnidos, la morfología completa —particularmente los patrones de calcificación interna de los centros vertebrales del gran tiburón blanco— ha sido comparada a la de los centros vertebrales bien preservados de los tiburones megadentados, incluyendo a C. megalodon y C. angustidens. La aparente similitud morfológica apoya un parentesco cercano de las especies megadentadas fósiles con los actuales grandes tiburones blancos.[27][81]
Con respecto a los orígenes del gran tiburón blanco, Gottfried y Fordyce han señalado que algunos fósiles de grandes tiburones blancos tienen cerca de 16 millones de años y preceden a los fósiles de la transición al Plioceno (hace 5.3 millones de años).[27] Adicionalmente, los registros del Oligoceno de C. megalodon,[19][27] contradicen la sugerencia de que Carcharocles chubutensis sea el ancestro inmediato de C. megalodon. Estos registros también indican que C. megalodon en realidad coexistió con Carcharocles angustidens.[27] Por lo tanto, los partidarios del género Carcharodon para C. megalodon afirman que los tiburones megadentados extintos deben ser incluidos en el género Carcharodon.[27]
Algunos paleontólogos argumentan que el género Otodus debería ser usado para los tiburones incluidos en el linaje Carcharocles y que dicho género debería ser descartado.[5]
Al presente, varios de los partidarios del género Carcharocles para C. megalodon (como Catalina Pimiento, Dana J. Ehret, Bruce J. MacFadden y Gordon Hubbell) aceptan que ambas especies pertenecen al orden Lamniformes, y que en ausencia de miembros vivientes de la familia Otodontidae, el gran tiburón blanco es la mejor analogía ecológica para C. megalodon.[5]
Supervivencia actual
Aunque los expertos consideran que no hay ninguna evidencia disponible que indique que el megalodón no está extinto, la idea de que una población superviviente pudiese seguir existiendo en la actualidad parece haberse asentado en el imaginario público.
La idea de su supervivencia reciente se inspira usualmente en el descubrimiento de un diente de C. megalodon por miembros del barco HMS Challenger en 1872, el cual algunos creyeron que sólo tenía 10 000 años.[82] Sin embargo, este diente se ha reexaminado, y los hallazgos indican que su edad es imprecisa.[82] Los análisis de dos dientes de megalodón encontrados por el HMS Challenger dan una fecha de entre 10 000 y 15 000 años de antigüedad, tiempo estimado para la deposición de la cantidad de manganeso acumulada sobre ellos. No obstante, es bastante probable que los dientes estuviesen ya fosilizados antes de que la costra de manganeso se desarrollase sobre ellos, por lo que su origen sería mucho más antiguo y no tendría nada de extraordinario. Varios autores opinan que las estimaciones recientes para estos dientes son imprecisas y que cualquier afirmación de la existencia de Carcharodon megalodon en tiempos post-pliocénicos es simplemente errónea, basada en metodología y experimentos obsoletos.[83] Roesch y otros recalcan el hecho de que los megalodontes eran probablemente animales costeros (como los tiburones blancos), por lo que una supervivencia en el fondo de los océanos sería extremadamente inverosímil.
El megalodón en la ficción
Desde que los restos de C. megalodon fueron descubiertos, han sido un objeto de fascinación. Ha sido retratado en varios trabajos de ficción, incluyendo películas y novelas, y continúa manteniendo su lugar entre los temas más populares de ficción que involucran monstruos marinos. Muchos de estos relatos de ficción postulan que al menos una población relicta de C. megalodon sobrevivió la extinción y acecha en las vastas profundidades del océano, y que algunos individuos lograron llegar a la superficie desde lo profundo, ya sea por la intervención humana o por medios naturales. La historia de Jim Shepard "Tedford and the Megalodon" es un buen ejemplo de esto.
Algunos trabajos de ficción (como Shark Attack 3: Megalodon, y el de Steve Alten la serie "Meg", e incluso Megalodon) representan incorrectamente a C. megalodon como una especie de más de 70 millones de años, estando viva en el tiempo de los dinosaurios. Los escritores de la película Shark Attack 3: Megalodon representaron esta idea al incluir una copia alterada de un libro del investigador de tiburones Richard Ellis, llamado "Great White Shark". La copia mostrada en el filme tenía varias páginas que no existen en el libro real. El autor demandó a la distribuidora de la película, Lions Gate Entertainment, pidiendo que se ponga fin a la distribución de la película, junto con 150 000 dólares por daños y perjuicios.[84] La novela de Steve Alten Meg: A Novel of Deep Terror es conocida por retratar este error con sus ilustraciones del prólogo y la portada en la que muestra a C. megalodon matando a un tiranosaurio en el mar. Y por último, la película de 2004, Megalodon, muestra el error en una escena de la película, diciendo uno de los protagonistas que el Megalodón se extinguío hace 65 millones de años.
Las películas Mega Shark Versus Giant Octopus (2009) y su secuela Mega Shark Versus Crocosaurus (2010) son parodias de películas de catástrofes con un tiburón gigante similar al megalodón en el papel principal.
En agosto de 2013, la cadena Discovery Channel inició su evento anual Shark Week con Megalodon: The Monster Shark Lives (en inglés "Megalodón: el tiburón monstruo vive"),[85] un controvertido[86][87] falso documental acerca de la supuesta existencia de esta criatura.[88] Este programa ha sido criticado debido a que es completamente ficticio; por ejemplo, todos los supuestos científicos representados en realidad eran actores.[89] La supuesta fotografía mostrada en el programa, tomada en diciembre de 1942, donde se podía ver a un Megalodón nadando junto a un submarino, se hizo viral rápidamente en Internet, pero se demostró que está manipulada a partir de imágenes de archivo de submarinos en el Atlántico durante la Segunda Guerra Mundial.[90] En 2014 Discovery volvió a emitir "The Monster Shark Lives," junto con un nuevo programa de una hora de duración, "Megalodon: The New Evidence" (megalodón: la nueva evidencia), y un programa dramatizado adicional titulado Shark of Darkness: Wrath of Submarine (Tiburón de la oscuridad: ira de Submarino), lo que terminó en un rechazo mayor de los medios de comunicación y de la comunidad científica.[91][92][93][94]
La película Megalodón (2018) ha sido el último medio de ficción donde se muestra al tiburón prehistórico, con la hipótesis de que el megalodón se escondía en las profundidades del océano Pacífico y que saldría a la superficie después de haber sido perturbado por la humanidad, y con el objetivo de devorar a Jonas Taylor (Jason Statham) y su tripulación.
Referencias
- Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (30 de marzo de 2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. doi:10.1111/jbi.12754.
- Pimiento, C.; Clements, C. F. (22 de octubre de 2014). «When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record». PLoS ONE 9 (10): e111086. doi:10.1371/journal.pone.0111086.
- Wroe, S.; Huber, D. R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T. L.; Cunningham, E.; Dean, M. N.; Summers, A. P. (2008). «Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?». Journal of Zoology 276 (4): 336-342. doi:10.1111/j.1469-7998.2008.00494.x.
- Pimiento, Catalina; Dana J. Ehret, Bruce J. MacFadden, and Gordon Hubbell (10 de mayo de 2010). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLoS One (Panama: PLoS.org) 5 (5): e10552. PMC 2866656. PMID 20479893. doi:10.1371/journal.pone.0010552. Consultado el 12 de mayo de 2010.
- Lambert, Olivier; Giovanni Bianucci, Klaas Post, Christian de Muizon, Rodolfo Salas-Gismondi, Mario Urbina and Jelle Reumer (1 de julio de 2010). «The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru». Nature (Peru) 466 (7302): 105-108. PMID 20596020. doi:10.1038/nature09067. Archivado desde el original el 20 de septiembre de 2011.
- Haven, Kendall (1997). 100 Greatest Science Discoveries of All Time. Libraries Unlimited. pp. 25-26. ISBN 1591582652.
- Steno, N. (1958) [1667]. Garboe, Axel, ed. The earliest geological treatise (1667) by Nicolas Steno (Niels Stensen). Translated from Canis Charcarie Dissectum Caput (en inglés). Londres: Macmillan and Company. p. 51.
- Bruner, John (1997). «The "Megatooth" shark, Carcharodon megalodon.» (en versión en inglés). Mundo Marino Revista Internacional de Vida Marina. Archivado desde el original el 16 de julio de 2015. Consultado el 22 de enero de 2012.
- Rudwick, M. J. S. (1987) [1976]. El significado de los fósiles. Episodios de la Historia de la Paleontología. Ciencias de la Naturaleza. Madrid: Hermann Blume. p. 347. ISBN 84-7214-371-6.
- Nyberg K. G., Ciampaglio C. N., Wray G. A. (2006). «Tracing the ancestry of the GREAT WHITE SHARK». Journal of Vertebrate Paleontology 26 (4): 806-814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2. Consultado el 25 de diciembre de 2007.
- Agassiz, Louis (1833-1843). Recherches sur les poissons fossiles ... / par Louis Agassiz.. Neuchatel :Petitpierre. p. 41. Consultado el 8 de septiembre de 2008.
- Klimley, Peter; Ainley, David (1996). Great White Sharks: The Biology of Carcharodon carcharias. Academic Press. ISBN 0124150314. Archivado desde el original el 12 de octubre de 2012. Consultado el 30 de agosto de 2011.
- Augilera, Orangel A.; Garcia, Luis; Cozzoul, Mario A. (2008). «Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation». Paläontologische Zeitschrift (Springer Berlin) 82 (2): 204-208. doi:10.1007/BF02988410. Archivado desde el original el 20 de marzo de 2020. Consultado el 30 de agosto de 2011.
- Narrated by: Robert Leigh (2009-04-27). «Monster Shark». Prehistoric Predators. National Geographic.
- Lawley, R. (1881). «Selache manzonii n. sp. – Dente Fossile délia Molassa Miocenica del Monte Titano (Repubblica di San Marino)» [Fossil tooth from Miocene Molasse from Monte Titano (Republic of San Marino)]. Atti della Società Toscana di Scienze Naturali (en italiano) 5: 167-172.
- Roesch, Ben (1998). «The Cryptozoology Review: A Critical Evaluation of the Supposed Contemporary Existence of Carcharocles Megalodon». Archivado desde el original el 21 de octubre de 2013.
- Bendix-Almgreen, Svend Erik (15 de noviembre de 1983). «Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn». Bulletin of the Geological Society of Denmark (Copenhagen: Geologisk Museum) 32: 1-32. Consultado el 5 de febrero de 2012.
- Renz, Mark (2002). Megalodon: Hunting the Hunter. PaleoPress. ISBN 0-9719477-0-8.
- Nieves-Rivera, Angel M.; Ruizyantin, Maria; Gottfried, Michael D. (2003). «New Record of the Lamnid Shark Carcharodon megalodon from the Middle Miocene of Puerto Rico». Caribbean Journal of Science 39: 223-227.
- Iturralde-Vinent, M.; G. Hubbell, and R. Rojas (1996). «Catalog Of Cuban Fossil Elasmobranchii (Paleocene--Pliocene) and Paleooceanographic Implications of Their Lower--Middle Miocene Occurrence». Boletín de la Sociedad Jamaicana de Geología (Cuba) 31: 7-21. Archivado desde el original el 14 de abril de 2010. Consultado el 10 de mayo de 2010.
- Donovan, Stephen; Gavin, Gunter (2001). «Fossil sharks from Jamaica». Bulletin of the Mizunami Fossil Museum 28: 211-215. Archivado desde el original el 31 de octubre de 2013.
- Fitzgerald, Erich (2004). «A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia». Memoirs of Museum Victoria (Australia: Museum Victoria) 61 (2): 183-208. ISSN 1447-2554. Archivado desde el original el 23 de agosto de 2008. Consultado el = marzo de 2010.
- Portell, Roger; Hubell, Gordon; Donovan, Stephen; Green, Jeremy; Harper, David; Pickerill, Ron (2008). Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles 44 (3). Caribbean Journal of Science. pp. 279-286.
- Hallan en Canarias restos del Megalodón, el tiburón más grande de la historia
- Hideo, Habe; Mastatoshi, Goto; Naotomo, Kaneko (22 de marzo de 2004). «Age of Carcharocles megalodon (Lamniformes: Otodontidae) : A review of the stratigraphic records». The Palaeontological Society of Japan (PSJ) (Japón) 75: 7-15. ISSN 0022-9202.
- Gottfried M. D., Fordyce R. E. (2001). «An Associated Specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) From the Late Oligocene of New Zealand, with comments on Carcharodon Interrelationships». Journal of Vertebrate Paleontology 21 (4): 730-739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2.
- Brown, Robin (2008). Florida's Fossils. Pineapple Press. ISBN 978-1-56164-409-4.
- Helfman, Gene; Collette, Bruce; Facey, Douglas (1997). The diversity of fishes. Wiley Blackwell. ISBN 978-0-8654-2256-8.
- Randall, John (julio de 1973). «Size of the Great White Shark (Carcharodon)». Science Magazine: 169-170.
- Kowinsky, Jayson (2002). «The Size of Megalodons». Consultado el 12 de enero de 2008.
- Shimada, Kenshu (5 de noviembre de 2002). «The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae)». Journal of Fossil Research (Japón) 35 (2): 28-33. ISSN 0387-1924. Archivado desde el original el 13 de marzo de 2012. Consultado el 29 de abril de 2011.
- Schembri, Patrick (1994). «Malta's Natural Heritage». Natural Heritage. in (Malta: University of Malta): 105-124. Consultado el marzo de 2010.
- Papson, Stephen (1992), «Copyright: Cross the Fin Line of Terror», Journal of American Culture 15: 67-81.
- Compagno, Leonard J. V. (2002). Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Rome: Food & Agriculture Organization of the United Nations. p. 97. ISBN 9251045437.
- Herskowitz, David; Peter Wiggins (21 de diciembre de 2010). «Huge Shark Jaw» (PDF). Natural History. Heritage Auctions. Consultado el 30 de abril de 2011.
- Anderson, Philip; Westneat, Mark (22 207 de febrero de 2007). «Feeding mechanics and bite force modelling of the skull of Dunkleosteus terrelli, an ancient apex predator». Royal Society 3 (1): 77-80. doi:10.1098/rsbl.2006.0569Bot inserted parameter. Either remove it; or change its value to "." for the cite to end in a ".", as necessary.
- Riordon, James (junio de 1999). «Hell's teeth». NewScientist Magazine (2190): 32.
- Andres, Lutz (2002). «C. megalodon — Megatooth Shark, Carcharodon versus Carcharocles». Consultado el 16 de enero de 2008.
- Arnold, Caroline (2000). Giant Shark: Megalodon, Prehistoric Super Predator. Houghton Mifflin. pp. 18-19. ISBN 9780395914199.
- Gillette, Lynett. «Winds of Change». San Diego Natural History Museum. Archivado desde el original el 4 de enero de 2011. Consultado el 25 de septiembre de 2009.
- Aguilera O., Aguilera E. R. D. (2004). «Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage». Caribbean Journal of Science 40 (3): 362-368.
- Collareta, A.; Lambert, O.; Landini, W.; Di Celma, C.; Malinverno, E.; Varas-Malca, R.; Urbina, M.; Bianucci, G. «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. doi:10.1016/j.palaeo.2017.01.001.
- «Fact File: Odobenocetops». BBC. Consultado el 21 de marzo de 2008.
- Morgan, Gary S. (agosto de 1994). «Whither the giant white shark?». Paleontology Topics. Paleontological Research Institution.
- «MEGALODON». Fossil Farm Museum Of The Fingerlakes. Archivado desde el original el 5 de agosto de 2010. Consultado el 1 de julio de 2010.
- deGruy, Michael (2006). Perfect Shark (TV-Series). Reino Unido: BBC.
- Godfrey, Stephen (abril de 2004). «The Ecphora: Fascinating Fossil Finds» (PDF). Paleontology Topics. Calvert Marine Museum. Archivado desde el original el 19 de septiembre de 2010. Consultado el 21 de febrero de 2010.
- Kehe, Andy. «Bone Apetite». Archivado desde el original el 18 de mayo de 2008. Consultado el 17 de febrero de 2011.
- Godfrey, Stephen (marzo de 2007). «The Ecphora: Shark-Bitten Sea Cow Rib» (PDF). Paleontology Topics. Calvert Marine Museum. Archivado desde el original el 20 de octubre de 2010. Consultado el 27 de marzo de 2010.
- Compagno, Leonard J. V. (mayo de 1989). «Copyright: Alternative life-history styles of cartilaginous fishes in time and space». Environmental Biology of Fishes 28: 33-75. doi:10.1007/BF00751027
- Ferretti, Francesco; Boris Worm, Gregory L. Britten, Michael R. Heithaus, and Heike K. Lotze1 (agosto de 2010). «Patterns and ecosystem consequences of shark declines in the ocean». Ecology Letters (Blackwell Publishing Ltd) 13 (8): 1055-1071. PMID 20528897. doi:10.1111/j.1461-0248.2010.01489.x. Archivado desde el original el 24 de mayo de 2011. Consultado el 19 de febrero de 2011.
- «New Leviathan Whale Was Prehistoric "Jaws"?». National Geographic Daily News (Peru: National Geographic). 30 de junio de 2010. Consultado el -3 de julio de 2010.
- Bianucci, Giovanni; Walter, Landini (8 de septiembre de 2006). «Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy». Zoological Journal of the Linnean Society 148 (1): 103-131. doi:10.1111/j.1096-3642.2006.00228.x
- «Ancient monster whale more fearsome than Moby Dick». NewScientist. Consultado el 30 de junio de 2010.
- Tanke, Darren; Currie, Philip (diciembre de 1998). Head-Biting Behaviour in Theropod Dinosaurs: Paleopathological Evidence. Gaia 15. p. 168. ISSN 0871-5424.
- Landini, W.; Altamirano-Sera, A.; Collareta, A.; Di Celma, C.; Urbina, M.; Bianucci, G. (2017). «The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru)». Journal of South American Earth Sciences 73: 168-190. doi:10.1016/j.jsames.2016.12.010.
- Deméré, Thomas A.; Berta, Annalisa; McGowen, Michael R. (2005). «The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes». Journal of Mammalian Evolution 12 (1/2): 99-143. doi:10.1007/s10914-005-6944-3.
- Identifican en Canarias fósiles de 'megalodón', el tiburón más grande que ha existido
- «Details of ancient shark attack preserved in fossil whale bone». Smithsonian. Consultado el 12 de noviembre de 2011.
- Domning, Daryl P. (1 de febrero de 2001). «Sirenians, seagrasses, and Cenozoic ecological change in the Caribbean». Palaeogeography, Palaeoclimatology, Palaeoecology (USA: ELSEVIER) 166 (1-2): 27-50. doi:10.1016/S0031-0182(00)00200-5. Archivado desde el original el 16 de agosto de 2011. Consultado el 26 de abril de 2011.
- «How the Isthmus of Panama Put Ice in the Arctic». 22 de marzo de 2004. Consultado el 20 de diciembre de 2008.
- «Pliocene epoch». Consultado el 16 de enero de 2008.
- «Pleistocene epoch». Archivado desde el original el 7 de febrero de 2010. Consultado el 16 de enero de 2008.
- Antunes, Miguel Telles; Ausenda Cáceres Balbino (marzo de 2010). «The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic». Revista Española de Paleontología (Portugal) 25 (1): 1-6. ISSN 0213-6937. Consultado el 19 de febrero de 2011.
- Reilly, Michael (29 de septiembre de 2009). «Prehistoric Shark Nursery Spawned Giants». USA: Discovery News. Consultado el <31 de marzo de 2010.
- Dooly A.C, Nicholas C.F, Luo Z.X (2004). «The Earliest known member of the rorqual—gray whale clade (Mammalia, Cetacea)». Journal of Vertebrate Paleontology 24 (2): 453-463. doi:10.1671/2401.
- Fordyce, R. Ewan; Patrick G. Quilty, and James Daniels (2002). «Australodelphis mirus, a bizarre new toothless ziphiid-like fossil dolphin (Cetacea: Delphinidae) from the Pliocene of Vestfold Hills, East Antarctica». Antarctic Science (Cambridge University Press) 14 (1): 37-54. doi:10.1017/S0954102002000561.
- Allmon, Warren D.; Steven D. Emslie, Douglas S. Jones, and Gary S. Morgan (2006). «Late Neogene Oceanographic Change along Florida's West Coast: Evidence and Mechanisms». The Journal of Geology (USA: The University of Chicago) 104 (2): 143-162. Bibcode:1996JG....104..143A. doi:10.1086/629811.
- Heyning, John; Dahlheim, Marilyn (15 de enero de 1988). «Orcinus orca». Mammalian Species 304: 1-9. Consultado el 4 de febrero de 2012.
- Turner, Pamela S. (Oct/Nov 2004). «Showdown at Sea: What happens when great white sharks go fin-to-fin with killer whales?». National Wildlife (National Wildlife Federation) 42 (6). Archivado desde el original el 25 de noviembre de 2009. Consultado el 22 de noviembre de 2009.
- http://www.upi.com/Science_News/2016/03/31/Megalodon-shark-was-outcompeted-for-shrinking-food-supply/6621459433501/
- Slater, G. J.; Goldbogen, J. A.; Pyenson, N. D. (24 de mayo de 2017). «Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics». Proceedings of the Royal Society B: Biological Sciences 284 (1855): 20170546. doi:10.1098/rspb.2017.0546.
- Ehret D. J., Hubbell G., Macfadden B. J. (2009). «Exceptional preservation of the white shark Carcharodon from the early Pliocene of Peru». Journal of Vertebrate Paleontology 29 (1): 1-13. doi:10.1671/039.029.0113.
- Siverson, Mikael; Johan Lindgren, Michael G. Newbrey, Peter Cederström und Todd D. Cook (2013). «Late Cretaceous (Cenomanian-Campanian) mid-palaeolatitude sharks of Cretalamna appendiculata type» (PDF). Acta Palaeontologica Polonica: 2. doi:10.4202/app.2012.0137. Archivado desde [ el original] el 29 de noviembre de 2015.
- Glickman, L. S. (1964). Akuly paleogena i ich stratigrafičeskoe značenie [Sharks of Paleogene and their stratigraphic significance] (en ruso). Moscow, USSR: Nauka Press. p. 229.
- Cappetta, H. (2006). «Mesozoic and Cenozoic Elasmobranchii». Handbook of Paleoichthyology. 3B. München Pfeil. ISBN 978-3-89937-046-1. OCLC 829906016.
- «New large prehistoric shark discovered». ScienceDaily. DePaul University. 3 de octubre de 2016. Consultado el 3 de octubre de 2016.
- Shimada, K.; Chandler, R. E.; Lam, O. L. T.; Tanaka, T.; Ward, D. J. (3 de octubre de 2016). «A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade». Historical Biology: 1-11. doi:10.1080/08912963.2016.1236795.
- Dell-Amore, Christine (2009). «Most Complete Great White Fossil Yet». Consultado el 27 de enero de 2012.
- Godfrey, Stephen (11 de noviembre de 2006). «The Geology and Paleontology of Calvert Cliffs» (PDF). Paleontology Topics. The Ecphora Miscellaneous Publications. Archivado desde el original el 19 de septiembre de 2010. Consultado el 2 de noviembre de 2009.
- «Mega Jaws». MonsterQuest. Episodio 7. Temporada 3. March 18, 2009. Archivado del original el 9 de febrero de 2010. https://web.archive.org/web/20100209160026/http://www.history.com/genericContent.do?id=62124.
- https://web.archive.org/web/20041016122428/http://www.ncf.carleton.ca/~bz050/megalodon.html
- Boniello, Kathianne (12 de julio de 2009). «Shark Film has Writer Biting Mad». New York Post. Archivado desde el original el 3 de julio de 2012.
- «Shark Week 'Megalodon: The Monster Shark Lives' Tries To Prove Existence Of Prehistoric Shark (VIDEO)». Huff Post Green. 5 de agosto de 2013. Consultado el 11 de agosto de 2013.
- Wilcox, Christie (5 de agosto de 2013). «Shark Week Jumps The Shark: An Open Letter To Discovery Communications». Discover Magazine. Archivado desde el original el 10 de septiembre de 2018. Consultado el 11 de agosto de 2013.
- Wilcox, Christie (9 de agosto de 2013). «Discovery’s Megalodon Defense? ‘We Don’t Know,’ Or ‘We Don’t Care’». Discover Magazine. Archivado desde el original el 16 de septiembre de 2018. Consultado el 11 de agosto de 2013.
- Juzwiak, Rich (5 de agosto de 2013). «Shark Week Opens With Fake Megalodon Documentary». gawker.com. Consultado el 11 de agosto de 2013.
- «Shark Week's dark side: After fake documentary controversy, Discovery doubles down on its lies». OregonLive.com. Consultado el 6 de agosto de 2014.
- «El supuesto Megalodón vivo fotografiado junto a un submarino».
- «Shark Week is once again making things up». vox.com. Consultado el 16 de agosto de 2014.
- «Discovery Revisits Megalodon For Shark Week 2014 Despite Backlash». inquisitr.com. Consultado el 16 de agosto de 2014.
- «Shark Week Is Lying Again About Megalodon Sharks». slate.com. Consultado el 16 de agosto de 2014.
- «Sorry, Fans. Discovery Has Jumped the Shark Week.». nytimes.com. Consultado el 16 de agosto de 2014.
{kind=link}
Bibliografía
- Agassiz, L. 1833-1844. Recherches sur les Poissons Fossiles, V. 1-5. Imprimerie de Petitpierre, Neuchâtel, Suisse.
- Bretton, W. Kent (1994). Fossil Sharks of the Chesapeake Bay Region. Egan Rees & Boyer, Inc. 146 páginas. ISBN 1-881620-01-8
- Dickson, K. A.; y J. B. Graham (2004). «Evolution and consequences of endothermy in fishes». Physiological and Biochemical Zoology 77 (6): 998-1018. PMID 15674772. doi:10.1086/423743.
Enlaces externos
 Wikispecies tiene un artículo sobre Carcharocles megalodon.
Wikispecies tiene un artículo sobre Carcharocles megalodon. Wikimedia Commons alberga una categoría multimedia sobre Carcharocles megalodon.
Wikimedia Commons alberga una categoría multimedia sobre Carcharocles megalodon.- Ferrón, Humberto (2019) «Megalodón, un tiburón extinto de sangre caliente».¡Fundamental!, 32: 1–46
- Megalodón, el coloso oceánico
- ¿Carcharodon o Carcharocles?
- Una evaluación crítica de la supuesta existencia contemporánea de Carcharodon megalodon, por Ben S. Roesch
- Reconstruyendo al Megalodon
- Ficha del megalodonte en la BBC, con reconstrucciones 3-D y vídeo
- Dykens, M.; Gillette, L. «SDNHM Fossil Field Guide: Carcharodon megalodon, Giant "Mega-Tooth" Shark». Archivado desde el original el 4 de enero de 2012. Consultado el 29 de abril de 2012.
- Museo de dientes fósiles hallados en Georgia (EE. UU.)
- «Buceando en busca de los monstruos marinos del pasado». Consultado el 22 de julio de 2010.
- La mordida de un antiguo tiburón es más poderosa que la de un T. Rex en LiveScience (en inglés)
- Guía de campo de fósiles, Carcharocles megalodon del Museo de Historia Natural de San Diego (en inglés)
- La reconstrucción de Megalodón de la BBC, con imágenes y video (en inglés)
- Información sobre el Prehistórico Megalodón (en inglés)
- Hechos sobre Megalodón: el mayor tiburón que haya vivido (en inglés)
- La mayor reconstrucción de la mandíbula del megalodón en el mundo del Museo de Naturaleza y Ciencia (en inglés)
- Historia de tiburones presentando a Megalodón con una demostración de la mordida del tiburón gigante por el Dr. Chuck Ciampaglio (en inglés)
- Jurassic Shark (en inglés)
- Imagen en 3-D del gran tiburón blanco comparado al Megalodón (en inglés)
- Tiburones prehistóricos mataban ballenas con una sola mordida por Steve Farrar, corresponsal de ciencia (en inglés)
- Portal del Océano: ilustración comparativa del tamaño de Megalodón por la Institución Smithsoniana (en inglés)
- Restos fósiles de un gran cachalote con marcas de mordida de C. megalodon (un ejemplo de las relaciones depredador-presa entre C. megalodon y los cachalotes) (en inglés)
{kind=link}
{kind=link}
Videos paleontológicos
NOTA: se necesita Flash Player para ver el siguiente contenido:
- Un videoclip del programa Perfect Shark (2006) de la BBC (Presenta evidencia fósil de la depredación de Megalodón) (en inglés)
- Galería de videos con el Megalodón en Discovery Channel (en inglés)
- El paleontólogo Mark Renz muestra un gran diente de Megalodón (uno de los mayores descubiertos) en YouTube. (en inglés)
- Comparación de tamaño animada del Megalodón con el gran tiburón blanco, un humano y un bus escolar del Museo de Ciencias Naturales de Carolina del Norte en YouTube.Museo de Ciencias Naturales de Carolina del Norte en YouTube.. (en inglés)
- Washington DC prehistórico: Megatiburón de Discovery Channel (muestra las estrategias de ataque del Megalodón) (en inglés)
- Especial de la Semana del Tiburón sobre Megalodón con Pat McCarthy y John Babiarz en YouTube. con comentarios sobre su extinción. (en inglés)
- Dientes fósiles de Megalodón muestran evidencia de un criadero de tiburones de diez millones de años en YouTube. (en inglés)
- Información sobre el Megalodón en YouTube. (presenta Dana Ehret) (en inglés)