Complejo mayor de histocompatibilidad
El complejo mayor de histocompatibilidad o CMH (también CPH por complejo principal de histocompatibilidad o MHC, acrónimo para el inglés de major histocompatibility complex), es una familia de genes hallados en todos los vertebrados y ubicados en el brazo corto del cromosoma 6 en humanos, cuya función es la codificación de moléculas (glucoproteínas) denominadas antígenos leucocitarios humanos o antígenos de histocompatibilidad, que participan en la presentación de antígenos a los linfocitos T permitiendo la activación de procesos críticos en la generación de la respuesta inmunitaria. En general, el CMH permite distinguir lo propio de lo extraño.[1][2][3]
En humanos, el CMH se denominó primeramente «complejo HLA» (del acrónimo inglés para human leucocytic antigen), porque las moléculas del CMH se descubrieron como antígenos que diferencian los leucocitos de distintas personas y que producen la respuesta inmunitaria del receptor en trasplantes, produciendo el rechazo.[4]
El CMH es la zona del genoma humano más variable y contiene numerosos genes funcionales, caracterizados por un gran polimorfismo. Los productos de estos genes son mencionados por muchos autores también como CMH o bien HLA.[1]
Historia
El CMH fue descubierto en la década de 1950. Previamente, en el año 1900 el campo de la inmunogenética se abrió con el descubrimiento de los grupos sanguíneos, seguido por el descubrimiento del factor Rh en 1940.[5]
En 1936, Peter Gorer fue el primero que describió un complejo de histocompatibilidad en ratones. Su investigación fue seguida por George Snell quién planteó que el rechazo de tejidos por el ratón era debido a incompatibilidad en algunos antígenos. Este CMH fue denominado «H-2» (en honor al antígeno 2 descrito por Gorer) y es similar al CMH de varias especies.[3][5]
En 1952, Jean Dusset planteó la hipótesis de que un sistema de histocompatibilidad similar al de los glóbulos rojos debía estar presente en los glóbulos blancos. Logró demostrarlo con la aglutinación de leucocitos en el suero de un paciente politransfundido. Su trabajo al respecto le valió el Premio Nobel en 1958.[1][5]
La primera identificación concreta de un producto del CMH, fue hecha en el año 1958, mediante el estudio de suero reactivo. Se identificó el antígeno MAC, correspondiente al actual HLA-A2. En los años 1960, el polimorfismo fue confirmado con el trabajo de Jon van Rod, Rose Payne y Walter Bodmer, quienes describieron los antígenos 4a y 4b (Bw4 y Bw6), y los HLA-A2 y HLA-A3 en un estudio de mujeres multíparas.[5]
En el año 1964, se inició un esfuerzo internacional para la caracterizaciòn del complejo, denominado Taller Internacional de Histocompatibilidad (IHW, acrónimo del inglés International Histocompatibility Workshop). Se definió entonces el área del cromosoma 6 donde se codifican los HLA A, B y C, que por entonces se creía que solo era expresado por leucocitos (por eso el nombre HLA: human leucocytic antigen)); también se mapeó el sistema del complemento como proveniente de la misma zona. En los años 1970, se identificaron los HLA de clase II y luego, con los avances de la biología molecular, la investigación se centró a nivel de genes más que en sus productos.[3][5]
En el año 1967 se ocupó por primera vez el término haplotipo en relación con el CMH. En el año 1969, Baruj Benacerraf demostró que el CMH no solo era el causante de la aloreactividad, sino que permitía activar la respuesta inmune en contra de un antígeno en particular. Este descubrimiento le significó obtener el premio Nobel de medicina en 1980.[5]
En 1987, Bjorkman logró elucidar la estructura del HLA-A2 y también de las moléculas de clase II. El año 1999 se logró definir la secuencia de nuclétidos del CMH, mediante los esfuerzos combinados del Consorcio de Secuenciación del CMH bajo la dirección de Stephen Beck, Daniel Geraghty, Hidetoshi Inoko y Lee Rowen.[5]
Entre los años 1980 y 2000 el conocimiento de los alelos presentes en el CMH pasó de unas pocas decenas, con varios millones de posibles combinaciones alotípicas a alrededor de 15 000 alelos. La región HLA-B se convirtió en la región genética más polimórfica del genoma humano, seguida de la región del HLA-A. La región de genes MHC contiene una densidad y diversidad de genes extremadamente alta; la variación genética dentro de esta región juega un papel vital en la susceptibilidad a las enfermedades autoinmunes, infecciosas, y otras enfermedades.[5]
Organización de los genes y regiones del CMH
El análisis comparativo de la organización de la región MHC entre especies muy alejadas ha revelado la presencia de reorganizaciones dentro de la región específicas de cada línea evolutiva y cambios en la complejidad de los genes.
La región MHC es la región más densa en genes y más polimórfica del genoma de los mamíferos, crítica para la inmunidad y para el éxito reproductivo.
En especies no humanas
La estructura de la región MHC se conoce al menos en siete especies de mamíferos euterios (placentarios), dos de aves, cinco peces teleósteos y en los tiburones. Hay grandes diferencias en la organización de la región MHC entre los mamíferos euterios y los no mamíferos. En euterios, la región está ordenada a lo largo del cromosoma en las regiones clase I-II-III, es muy densa en genes y ocupa una zona extensa. En los no mamíferos, la región MHC generalmente contiene menos genes y las regiones Clase I y II son adyacentes, a excepción de los teleósteos, donde las dos regiones no están ligadas. De las regiones MHC secuenciadas completamente, la menos compleja es la del pollo, que contiene solo 19 genes en 92 kb.[6]
La región MHC en los marsupiales Monodelphis doméstica (Didelphimorphia gris de cola corta) está flanqueada por los marcadores genéticos MOG y COL11A2, comprende 3.95 Mb y contiene 114 genes, 87 compartidos con los humanos.[6]
En humanos
En humanos, el CMH está constituido por al menos de 4 000 000 y que podrían ser hasta 7 000 000 de pares de bases que codifican más de 200 genes ubicados en el brazo corto del cromosoma 6.[3][4]
EL CMH está dividido en 2 regiones de genes: la más cercana al centrómero contiene los genes de clase II, con los loci HLA–DRA, DRB, DQA, DQB, DPA, DPB, DNA, DMA, DMB y DOB; la región más cercan al telómero, contiene los genes de la clase I, con los loci HLA–A, B y C; entre ambas se ubican los genes de algunas proteínas del sistema de complemento y otros relacionados con la respuesta inmune.[1][3]
La región del brazo corto del cromosoma 6 que contiene los genes del CMH, posee la información de:
- ciertas glucoproteínas de la membrana plasmática involucradas en los mecanismos de presentación y procesamiento de antígenos a los linfocitos T: se agrupan en los genes de clase II (que codifican las proteínas CMH de clase II) y los genes de clase I (que codifican las proteínas CMH de clase I)
- citocinas y proteínas del sistema del complemento, importantes en la respuesta inmunológica, pero que no tienen nada que ver con los genes del CMH; estos genes se agrupan en la clase III.
Ambos tipos de moléculas participan en la respuesta inmunitaria, que permite la identificación de las moléculas propias y de las extrañas (invasoras), para eliminar estas últimas mediante diferentes mecanismos.

CMH de clase I
En euterios, la región CMH de clase I contiene un conjunto de genes metópicos cuya presencia y orden está conservada entre especies. Estas moléculas se expresan en todas las células humanas, excepto en los glóbulos rojos, las células germinales, las células de los embriones pre-implantación y el sincitiotrofoblasto (tejido embrionario, no presente en la vida extrauterina).[7] Algunas células, como las neuronas, los monocitos y los hepatocitos, presentan niveles bajos de moléculas del CMH de clase I (menos de 103 por célula).[8]
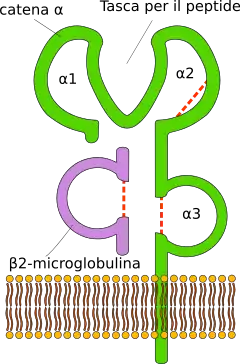
Los genes CMH de clase I se ubican en la zona más distal (cerca del telómero) y son el HLA-A, el HLA-B y el HLA-C. Son altamente polimórficos: se han descrito alrededor de 3000 alelos para cada uno. Junto a estos genes «clásicos» (o 1a) se ubican otros denominados «no clásicos» (o 1b): HLA-E, HLA-F, HLA-G, HFE y los MIC (acrónimo de cadena relacionadas con CMH de clase I, en inglés, MHC class I Chain-related): MICA y MICB. Excepto los MIC, todos estos genes codifican glucoproteínas, expresándose de manera codominante como heterodímeros en la membrana de cada célula nucleada, unido en forma no covalente con la cadena β2 microglobulina codificada por un gen del cromosoma 15.[1][2][4]
Sus productos tienen estructura de inmunoglobulina: presentan una cadena pesada tipo α que se subdivide en tres regiones: α1, α2 y α3. Estas tres regiones están expuestas al espacio extracelular y están unidas a la membrana de la célula mediante una región transmembrana. La cadena α está siempre asociada a una molécula de microglobulina β2, que está codificada por una región independiente en el cromosoma 15.
La principal función de los productos génicos de la clase I es la presentación de péptidos antigénicos intracelulares a los linfocitos T citotóxicos (CD8+). El péptido antigénico se aloja en una hendidura que se forma entre las regiones α1 y α2 de la cadena pesada, mientras el reconocimiento del MHC-I por parte del linfocito T citotóxico se hace en la cadena α3. En esta hendidura conformada por las regiones α1 y α2, se presentan péptidos de entre 8 y 11 aminoácidos, razón por la cual la presentación del péptido antigénico debe pasar por un proceso de fragmentación dentro de la propia célula que lo expresa.
En humanos, existen muchos isotipos (genes diferentes) de las moléculas de Clase-I, que pueden agruparse en:
- "clásicas", cuya función consiste en la presentación de antígenos a los linfocitos T CD8: dentro de este grupo tenemos HLA-A, HLA-B y HLA-C.
- "no clásicas" (también llamadas MHC clase IB), con funciones especializadas: no presentan antígenos a los linfocitos T, sino que se unen a receptores inhibidores de las células NK; dentro de este grupo se encuentran HLA-E, HLA-F, HLA-G. Por eso las proteínas HLA-G se denominan inmunosupresoras y se expresan en el citotrofoblasto del feto. Se piensa que esta expresión evita que el feto sea rechazado como un trasplante .
CMH de clase II
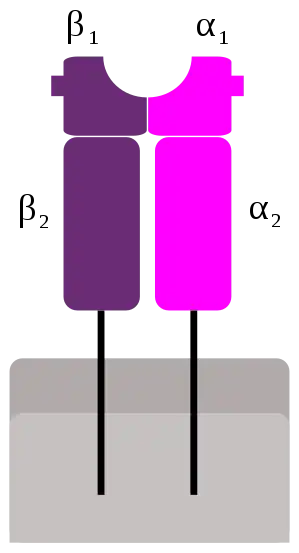
Los genes del CMH de clase II se encuentran en la región más cercana al centrómero, y codifican las dos cadenas del heterodímero funcional: HLA-DR, HLA-DQ, HLA-DP, HLA-DM, y HLA-DO. Inicialmente fueron denominados genes Ir por estar involucrados en la respuesta inmune (del acrónimo inglés para immune response).[1]
Estos genes codifican glucoproteínas con estructura de inmunoglobulina, pero en este caso el complejo funcional está formado por dos cadenas, una α y una β (cada una de ellas con dos dominios, α1 y α2, β1 y β2). Cada una de las cadenas está unida a la membrana por una región transmembrana, y ambas cadenas están enfrentadas, con los dominios 1 y 2 contiguos, en el exterior celular.
Estas moléculas se expresan de forma más restringidas que las de Clase I en las células presentadoras de antígeno, tales como células dendríticas, macrófagos, células de Langerhans, células de Kupffer, y en los linfocitos B.[1]
Las células presentadoras de antígenos presentan péptidos antigénicos extracelulares procesados a los linfocitos T cooperadores (CD4+). El péptido antigénico se aloja en una hendidura formada por los dominios α1 y β1, mientras el reconocimiento del MHC-II por parte del linfocito T cooperador se hace en la cadena β2. En esta hendidura conformada por las regiones α1 y β1, se presentan péptidos de entre 12 y 16 aminoácidos.
Las moléculas CMH de clase II en humanos presentan 5-6 isotipos, y pueden agruparse en:
- "clásicas", que presentan péptidos a los linfocitos T CD4; dentro de este grupo tenemos HLA-DP, HLA-DQ, HLA-DR;
- "no clásicas", accesorias, con funciones intracelulares (no están expuestas en la membrana celular, sino en membranas internas, de los lisosomas); normalmente, cargan los péptidos antigénicos sobre las moléculas MHC-II clásicas; en este grupo se incluyen HLA-DM y HLA-DO.
Además de las moléculas CMH de clase II, en la región Clase-II se encuentran genes que codifican moléculas procesadoras de antígenos, como TAP (por transporter associated with antigen processing) y Tapasin.
Región Intermedia
Entre las regiones ocupadas por los genes de clase I y de clase II, se encuentran numerosos genes que tienen importancia en la respuesta inmune pero no están relacionadas al CMH. Codifican varias proteínas que desempeñan funciones inmunitarias: componentes del sistema del complemento (como C2, C3 y factor B) y moléculas relacionadas con la inflamación (citoquinas como TNF-α, Linfotoxinas A y B) o proteínas de choque térmico.[1]
Polimorfismo del CMH
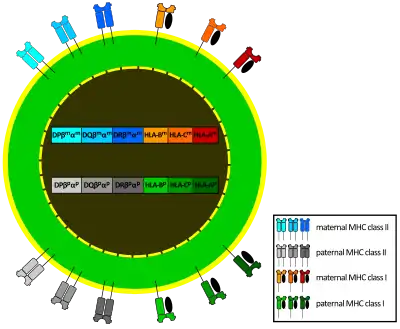
La región del CMH tiene dos características que dificultan a los patógenos la evasión selectiva de la respuesta inmunitaria: es poligénica y polimórfica, es decir, contiene varios genes diferentes de clase I y de clase II, y de cada uno existen múltiples variantes o alelos.[4]
Los genes del CMH se expresan de forma codominante. Esto quiere decir que los alelos heredados de ambos progenitores se expresan de forma equivalente:[4]
- Como existen tres pares genes que codifican la cadena α de la clase I, denominados en humanos HLA-A, HLA-B y HLA-C, y cada persona hereda un juego de cada progenitor, cualquier célula de un individuo podrá expresar hasta seis tipos diferentes de moléculas CMH de clase I.
- De forma parecida, en los loci de clase II, cada individuo hereda un par de genes HLA-DP (cada uno con DPA y DPB, que codifican las cadenas α y β respectivamente), un par de genes HLA-DQ (DQA y DQB, para las cadenas α y β), un gen HLA-DRα (DRA1) y uno o dos genes HLA-DRβ (DRB1 y DRB3, -4 o -5). Así, un individuo heterocigoto puede heredar 6 u 8 alelos de Clase-II, tres o cuatro de cada progenitor.
El juego de alelos presente en cada cromosoma se denomina haplotipo MHC. En humanos, cada alelo HLA recibe un número. Por ejemplo, para un individuo dado, su haplotipo puede ser HLA-A2, HLA-B5, HLA-DR3, etc. Cada individuo heterocigoto tendrá dos haplotipos MHC, uno en cada cromosoma (uno de origen paterno y otro de origen materno).
Los genes MHC son enormemente polimórficos, lo que significa que existen muchos alelos diferentes en los diferentes individuos de la población. El polimorfismo es tan grande que en una población mixta (no endogámica) no existen dos individuos que tengan exactamente el mismo juego de genes y moléculas MHC, excepto los gemelos idénticos.
Las regiones polimórficas de cada alelo se encuentran en la zona de contacto con el péptido que va a presentar al linfocito. Por esta razón, la zona de contacto de cada alelo de molécula MHC es muy variable, ya que los residuos polimórficos del MHC forman hendiduras específicas en las que las que solo pueden introducirse cierto tipo de residuo del péptido, lo cual impone un modo de unión muy preciso entre el péptido y la molécula MHC. Esto implica que cada variante de molécula MHC podrá unir específicamente solo aquellos péptidos que encajen adecuadamente en la hendidura de la molécula MHC, que es variable para cada alelo. De esta manera, las moléculas de MHC tienen una especificidad amplia para la unión de péptidos, puesto que cada molécula de MHC puede unir muchos, pero no todos los tipos de péptidos posibles. Esta es una característica esencial de las moléculas MHC: en un individuo concreto, bastan unas pocas moléculas diferentes para poder presentar una vasta variedad de péptidos.
Por otro lado, dentro de una población, la existencia de múltiples alelos asegura que siempre habrá algún individuo que posea una molécula de MHC capaz de cargar el péptido adecuado para reconocer un microbio concreto.La evolución del polimorfismo de MHC asegura que una población será capaz de defenderse frente a la enorme diversidad de microbios existentes, y que no sucumbirá ante la presencia de un nuevo patógeno o un patógeno mutado, porque al menos algunos individuos serán capaces de desarrollar una respuesta inmune adecuada para vencer al patógeno. Las variaciones en las secuencias de MHC (responsables del polimorfismo) resultan de la herencia de diferentes moléculas MHC, y no son inducidas por recombinación, como ocurre con los receptores de los antígenos.
Productos del CMH
Funciones de las moléculas del CMH de clase I y clase II
Ambos tipos de moléculas presentan péptidos antigénicos a los linfocitos T, responsables de la respuesta inmune específica para eliminar el patógeno responsable de la producción de dichos antígenos. Sin embargo, las moléculas MHC de Clase-I y II corresponden a dos vías diferentes de procesamiento de los antígenos, y se asocian con dos sistemas diferentes de defensa inmunitaria:
| Característica | Vía CMH de clase II | Vía CMH de clase I |
|---|---|---|
| Composición del complejo estable péptido-CMH | Cadenas polimórficas α y β, péptido unido a ambas | Cadena polimórfica α y microglobulina β2, péptido unido a cadena α |
| Tipos de células presentadoras de antígenos (APC) | Células dendríticas, fagocitos mononucleares, linfocitos B, algunas células endoteliales, epitelio del timo | Casi todas las células nucleadas |
| Linfocitos T capaces de responder | Linfocitos T cooperadores (CD4+) | Linfocitos T citotóxicos (CD8+) |
| Origen de las proteínas antigénicas | Proteínas presentes en endosomas o lisosomas (en su mayoría internalizadas del medio extracelular) | Proteínas citosólicas (en su mayor parte sintetizadas por la célula; pueden entrar también del exterior mediante fagosomas) |
| Enzimas responsables de la generación de péptidos | Proteasas de los endosomas y lisosomas (como la catepsina) | El proteasoma citosólico |
| Sitio de carga del péptido sobre la molécula del CMH | Compartimento vesicular especializado | Retículo endoplásmico |
| Moléculas implicadas en el transporte de los péptidos y carga sobre las moléculas del CMH | Cadena invariante, DM | TAP (transporter associated with antigen processing) |
Los linfocitos T de un individuo concreto presentan una propiedad denominada restricción del CMH: solo pueden detectar un antígeno si éste viene presentado por una molécula del CMH del mismo individuo. Esto se debe a que cada linfocito T tiene una especificidad dual: el receptor del linfocito T (denominado TCR por T cell receptor) reconoce algunos residuos del péptido y simultáneamente algunos residuos de la molécula del CMH que lo presenta. Esta propiedad es muy importante en el trasplante de órganos, e implica que, durante su desarrollo, los linfocitos T deben "aprender" a reconocer las moléculas MHC propias del individuo, mediante el proceso complejo de maduración y selección que tiene lugar en el timo.
Las moléculas del CMH solo pueden presentar péptidos, lo que implica que los linfocitos T, dado que solo pueden reconocer un antígeno si viene asociado a una molécula del CMH, solo pueden reaccionar ante antígenos de origen proteico (procedentes de microbios) y no a otro tipo de compuestos químicos (ni lípidos, ni ácidos nucleicos, ni azúcares). En cambio, la molécula CD1 de estructura similar a las moléculas del CMH de clase I (3 cadenas α y una β-microglobulina) presenta antígenos no peptídicos, generalmente lípidos, a linfocitos T. Cada molécula MHC puede presentar un único péptido cada vez, dado que la hendidura de la molécula solo tiene espacio para alojar un péptido. Sin embargo, una molécula del CMH dada tiene una especificidad amplia, porque puede presentar muchos péptidos diferentes (aunque no todos).

Las moléculas del CMH adquieren el péptido que presentan en el exterior de la membrana celular durante su propia biosíntesis, en el interior celular. Por tanto, los péptidos que presentan las moléculas del CMH provienen de microbios que están en el interior celular, y ésta es la razón por la cual los linfocitos T, que solo identifican péptidos cuando están asociados a moléculas del CMH, solo detectan microbios asociados a células y desencadenan una respuesta inmune contra microbios intracelulares.
Es importante resaltar que las moléculas del CMH de clase I adquieren péptidos que provienen de proteínas citosólicas, mientras que las moléculas del CMH de clase II adquieren péptidos de proteínas contenidas en vesículas intracelulares. Por ello, las moléculas del CMH de clase I presentarán péptidos propios, péptidos virales (sintetizados por la propia célula) o péptidos procedentes de microbios ingeridos en fagosomas. Las moléculas del CMH de clase II, por su parte, presentarán péptidos procedentes de microbios ingeridos en vesículas (este tipo de moléculas solo se expresan en células con capacidad fagocítica). Las moléculas del CMH solo se expresan de forma estable en la membrana celular si tienen un péptido cargado: la presencia del péptido estabiliza la estructura de las moléculas del CMH, las moléculas "vacías" se degradan en el interior celular. Las moléculas del CMH cargadas con un péptido pueden permanecer en la membrana durante días, el tiempo suficiente para asegurar que un linfocito T adecuado reconozca el complejo e inicie la respuesta inmunitaria.
En cada individuo, las moléculas del CMH pueden presentar tanto péptidos extraños (procedentes de patógenos) como péptidos procedentes de las proteínas propias del individuo. Esto implica que, en un momento dado, solo una pequeña proporción de las moléculas del CMH de una célula presentará un péptido extraño: la mayor parte de los péptidos que presente serán propios, dado que son más abundantes. Sin embargo, los linfocitos T son capaces de detectar un péptido presentado por solo el 0,1%-1% de las moléculas del CMH para iniciar una respuesta inmune.
Los péptidos propios, por otro lado, no pueden iniciar una respuesta inmune (excepto en los casos de las enfermedades autoinmunes), porque los linfocitos T específicos para los antígenos propios son destruidos o inactivados en el timo. Sin embargo, la presencia de péptidos propios asociados a las moléculas MHC es esencial para la función supervisora de los linfocitos T: estas células están constantemente patrullando el organismo, verificando la presencia de péptidos propios asociados a las moléculas del CMH y desencadenando una respuesta inmune en los casos raros en los que detectan un péptido extraño.
Las moléculas del CMH en el rechazo de trasplantes
Las moléculas del CMH se identificaron y nombraron precisamente por su papel en el rechazo de trasplantes entre ratones de diferentes cepas endogámicas. En humanos, las moléculas MHC son los antígenos de los leucocitos (HLA). Llevó más de 20 años comprender la función fisiológica de las moléculas del CMH en la presentación de péptidos a los linfocitos T.
Como se ha indicado antes, cada célula humana expresa 6 alelos del CMH de clase I (un alelo HLA-A, -B y -C de cada progenitor) y 6-8 alelos del CMH de clase 2 (uno HLA-DP y -DQ, y uno o dos de HLA-DR de cada progenitor, y algunas combinaciones de éstos). El polimorfismo de los genes del CMH es muy elevado: se estima que en la población hay al menos 350 alelos de los genes HLA-A, 620 alelos de HLA-B, 400 alelos de DR y 90 alelos de DQ. Como estos alelos pueden heredarse y expresarse en muchas combinaciones diferentes, cada individuo expresará probablemente algunas moléculas que serán diferentes de las moléculas de otro individuo, excepto en el caso de los gemelos idénticos. Todas las moléculas MHC pueden ser dianas del rechazo de trasplantes, aunque HLA-C y HLA-DP tienen un bajo polimorfismo, y probablemente tengan una importancia menor en los rechazos.
En el caso de un trasplante (de órganos o de células madre), las moléculas HLA funcionan como antígenos: pueden desencadenar una respuesta inmunitaria en el receptor, provocando el rechazo del trasplante. El reconocimiento de los antígenos del CMH en células de otro individuo es una de las respuestas inmunes más intensas que se conocen. La razón por la que los individuos reaccionan contra las moléculas del CMH de otro individuo se comprende bastante bien.
Durante el proceso de maduración de los linfocitos T, éstos son seleccionados en función de la capacidad de su TCR de reconocer débilmente complejos "péptido propio:CMH propio". Por ello, en principio, los linfocitos T no deberían reaccionar frente a un complejo "péptido extraño:MHC extraño", que es lo que aparecerá en las células trasplantadas. Sin embargo, parece que lo que ocurre es un tipo de reacción cruzada: los linfocitos T del individuo receptor pueden equivocarse, porque la molécula del CMH del donante es similar a la propia en la zona de unión al TCR (la zona variable del CMH se encuentra en la zona de unión al péptido que presentan). Por esta razón, los linfocitos del individuo receptor interpretan el complejo presente en las células del órgano trasplantado como "péptido extraño:MHC propio" y desencadenan una respuesta inmune contra el órgano "invasor", porque lo perciben de la misma manera que un tejido propio infectado o tumoral, pero con un número mucho más elevado de complejos capaces de iniciar una respuesta. El reconocimiento de la molécula de MHC extraña como propia por los linfocitos T se denomina alorreconocimiento.
Pueden producirse dos tipos de rechazo de trasplantes mediado por las moléculas del CHM o antígenos leucocitarios humanos (HLA):
- rechazo hiperagudo: se produce cuando el individuo receptor presenta anticuerpos anti-HLA preformados, antes del trasplante; esto puede deberse a la realización previa de transfusiones de sangre (que incluye linfocitos del donante, con moléculas HLA), a la generación de anti-HLA durante el embarazo (contra los HLA del padre presentes en el feto) y por la realización de un trasplante previo;
- rechazo humoral agudo y disfunción crónica del órgano trasplantado: se debe a la formación de anticuerpos anti-HLA en el receptor, contra las moléculas HLA presentes en las células endoteliales del trasplante.
En ambos casos, se produce una reacción inmune contra el órgano trasplantado, que puede generar lesiones en el mismo, lo que conlleva la pérdida de función, inmediata en el primer caso y progresiva en el segundo.
Por esta razón, es fundamental realizar una prueba de reacción cruzada entre células del donante y suero del receptor, para detectar la presencia de anticuerpos anti-HLA preformados en el receptor contra las moléculas HLA del donante, y evitar el rechazo hiperagudo. Normalmente, se verifica la compatibilidad de las moléculas HLA-A, -B y -DR: a medida que aumenta el número de incompatibilidades, la supervivencia a 5 años del trasplante disminuye. La compatibilidad total solo existe entre gemelos idénticos, pero en la actualidad existen bases de datos de donantes a nivel mundial que permiten optimizar la compatibilidad HLA entre un donante potencial y un receptor.
Exogamia y CMH
En ratones, ovejas, peces (como el espinoso), aves y humanos, hay estudios que indicarían que la selección de pareja tiene que ver con la disimilitud en el CMH y su relación con el olor de la eventual pareja. Hay alguna evidencia, especialmente en ratones y espinosos, y menos en humanos, respecto de que se prefiere el olor de individuos con diferentes CMH, favoreciendo la diversidad y así una mejor sistema inmunitario de los descendientes.[9][10][11]
En humanos, la elección de pareja está determinada por múltiples factores, tales como simetría facial, aspectos psicológicos, condición de la piel, y quizá el olor corporal. Los resultados de los estudios han demostrado cierta relación entre las preferencias de pareja y la disimilitud en el CMH. Esto traería como consecuencia una mayor variabilidad genética de los descendientes.[12][13]
Véase también
Bibliografía
- Jeffrey K. Actor. Elsevier’s integrated review immunology and microbiology. Segunda edición. Huston, Estados Unidos: Elsevier Health Sciences, 2011.
- Abbas, A. B. Basic Immunology. Functions and disorders of the immune system. Tercera Edición. 2009.
Referencias
- Goldberg, Anna Carla; Rizzo, Luiz Vicente (enero a marzo de 2015). «MHC structure and function – antigen presentation. Part 1» [Estructura y función del MHC – presentación de antígenos. Parte 1]. Einstein (São Paulo) (en inglés) (San Pablo, Brasil: Scielo) 13 (1): 153-156. ISSN 2317-6385. doi:10.1590/S1679-45082015RB3122. Consultado el 27 de octubre de 2017.
- Vega Robledo, Gloria Bertha (marzo a abril de 2009). «Complejo mayor de histocompatibilidad». Rev Fac Med UNAM (México: medigraphic) 52 (2): 86-88. Consultado el 27 de octubre de 2017.
- López–Martínez, Alondra; Chávez–Muñoz, Claudia; Granados, Julio (marzo a abril de 2005). «Función biológica del complejo principal de histocompatibilidad». Revista de investigación clínica (México) 57 (2): 132-141. ISSN 0034-8376. Consultado el 10 de julio de 2016.
- Janeway, C. A. Jr.; Travers, P.; Walport, M. (2001). «The major histocompatibility complex and its functions» [El complejo mayor de histocompatibilidad y sus funciones]. Immunobiology: The Immune System in Health and Disease [Inmunobiología: el sistema inmune en la salud y la enfermedad] (en inglés) (5 edición). Nueva York: Garland Science. Consultado el 27 de octubre de 2017.
- Vandiedonck, Claire; Knight, Julian C. (25 de mayo de 2009). «The human Major Histocompatibility Complex as a paradigm in genomics research» [El CMH como paradigma de la investigación genética]. Brief Funct Genomic Proteomic (en inglés) (Oxford, Reino Unido: PubMed Central) 8 (5): 379-94. PMID 19468039. doi:10.1093/bfgp/elp010. Consultado el 11 de julio de 2016.
- Belov, Katherine; Deakin, Janine E.; Papenfuss, Anthony T.; Baker, Michelle L.; Melman, Sandra D.; Siddle, Hannah V.; et al (marzo de 2006). «Reconstructing an Ancestral Mammalian Immune Supercomplex from a Marsupial Major Histocompatibility Complex» [Reconstruyendo un supercomplejo inmune mamífero ancestral a partir del complejo mayor de histocompatibilidad de los marsupiales]. PLoS Biol (en inglés) (Public Library of Science) 4 (3): e46. PMID 16435885. doi:10.1371/journal.pbio.0040046. Consultado el 27 de octubre de 2017.
- Comiskey, Martina; Warner, Carol M.; Schust, Danny J. (2000-2013). Landes Bioscience, ed. MHC Molecules of the Preimplantation Embryo and Trophoblast [Moléculas del CMH del embrión preimplantado y del trofoblasto]. Texas, Estados Unidos: Madame Curie Bioscience Database [Internet]. NBK6277.
- Berke, Gideon; Clark, William R. (2007). «Ch.1 Basic Immunology: A Primer». Killer lymphocytes [Linfocitos asesinos] (2 edición). Springer Science & Business Media. ISBN 9781402065637.
- O’Dwyera, Terence W.; Nevitta, Gabrielle A. (2009). «Individual odor recognition in procellariiform chicks: Potential role for the major histocompatibility complex» [Reconocimiento individual del olor en crías de Procellariiformes: rol potencial del complejo mayor de histocompatibilidad]. Annals of the New York Academy of Sciences (en inglés) (New York Academy of Sciences) 1170: 442-446. doi:10.1111/j.1749-6632.2009.03887.x. Consultado el 12 de julio de 2016.
- Milinski, Manfred; Griffiths, Siân; Wegner, Mathias; Reusch, Thorsten B. H.; Haas-Assenbaum, Annette; Boehm, Thomas (22 de marzo de 2005). «Mate choice decisions of stickleback females predictably modified by MHC peptide ligands» [Predictibilidad en la decisión de elección de pareja en hembras de espinoso modificada por ligandos peptídicos del CMH]. Proceedings of the National Academy of Sciences of the United States of America (en inglés) (The National Academy of Sciences) 102 (12): 4414-4418. PMID 15755811. doi:10.1073/pnas.0408264102. Consultado el 12 de julio de 2016.
- Penn, Dustin J.; Potts, Wayne K. (febrero de 1999). «The evolution of mating preferences and major histocompatibility complex genes» [La evolución de preferencias de emparejamiento y los genes del complejo mayor de histocompatibilidad]. American Naturalist (en inglés) (Utah, Estado Unidos) 153 (2): 145-164. Consultado el 12 de julio de 2016.
- Thompson, Patrycja (27 de enero de 2013). «Mismatched MHC and human mating choices» [CMH disímiles y elección de pareja en humanos]. Immpress magazine (en inglés). Consultado el 12 de julio de 2016.
- «Opposites Attract: How Genetics Influences Humans To Choose Their Mates» [Atracción de opuestos: como la genética influye en la elección de pareja de los humanos]. ScienceDaily (en inglés). 25 de mayo de 2009. Consultado el 12 de julio de 2016.
| Control de autoridades |
|
|---|
 Datos: Q423163
Datos: Q423163 Multimedia: Major histocompatibility complex / Q423163
Multimedia: Major histocompatibility complex / Q423163