Morfógeno
Un morfógeno es una sustancia que determina el patrón del desarrollo tisular de acuerdo a su difusión en el medio.[1] En particular, controla el nivel de compromiso de varios tipos celulares dentro de un tejido en formación.
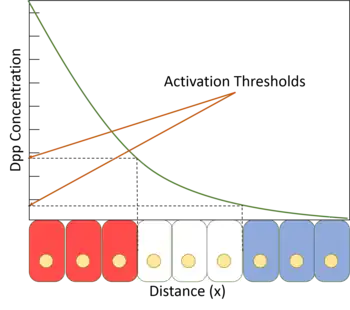
Su efecto se da por la acción directa de la sustancia que se expande desde una fuente localizada, formando un gradiente de concentración a lo largo de un tejido en desarrollo, resultando en una segmentación del embrión y una posterior especialización del tejido.[2]
Historia
El término fue acuñado por Alan Turing en su libro La base química de la morfogénesis. En la cual predijo un mecanismo químico que rige la segmentación de los organismos multicelulares.[3]
El concepto comenzó su historia en la Biología del desarrollo con los trabajos de Thomas Hunt Morgan en la mosca de la fruta (Drosophila melanogaster) a inicio del siglo XX. Posteriormente en 1960, Lewis Wolpert refinó el concepto mediante el modelo de bandera francesa, donde se describe el mecanismo de los morfógenos para generar diferentes niveles de compromiso celular según su distancia a la fuente de emisión. Este modelo fue complementado por el genetista Peter Lawrence.
La bióloga alemana, Christiane Nusslein-Volhard descubrió el primer morfógeno: Bicoid. Un Factor de transcripción de Efecto Materno presente en el estadio sincitial del embrión de Drosophila. Este trabajo la hizo ganadora del Premio Nobel de Fisiología o Medicina de 1995. Posteriormente Gary Struhl y Stephen Cohen describieron como la molécula Decapenthaplegic actúa como morfógeno en estadios posteriores de Drosophila.
Desde ese entonces se han estudiado diferentes morfógenos y sus homólogos para todos los organismos.
Mecanismos
Véase: Diferenciación celular
Durante el desarrollo temprano, los gradientes morfogenéticos generan diferentes tipos celulares en un orden espacial diferente. El morfógeno se une a los dominios extracelulares de proteínas receptoras, que mediante la transducción de señales comunican el nivel de morfógeno al núcleo. Estas cascadas de señalización suelen terminar en la activación de Factores de Trascripción.[4]
La expresión de los genes nucleares está controlada por segmentos del ADN llamados enhancers sobre los que los factores de transcripción actúan directamente. Una vez unidos, el factor de transcripción activa o inhibe la transcripción del gen y de ese modo controla el nivel de expresión del producto génico (por lo general una proteína).

Los genes nucleares con un umbral de activación bajo requieran bajos niveles del morfógeno para ser regulados. En esto casos, los enhancers contienen lugares de unión de alta afinidad para el factor de transcripción. Los genes nucleares de umbral de activación elevado tienen por lo general menos lugares de unión, por lo que requieren niveles mucho más altos de actividad del factor de transcripción para ser regulados.
Estadio sincitial en Drosophila
Drosophila melanogaster tiene un sistema de desarrollo inusual ya que en los primeros trece ciclos de división del embrión ocurren dentro de un sincitio, en esencia, el embrión permanece como una sola célula con alrededor de 8.000 núcleos uniformemente distribuidos cerca de la membrana. Solo es hasta la decimocuarta división que membranas independientes se sitúan entre los núcleos y forman células independientes (Celularización).
Como resultado, en los embriones de Drosophila los factores de transcripción como Bicoid o Hunchback pueden actuar como morfógenos, puesto que pueden difundirse libremente entre los núcleos libres para producir suaves gradientes de concentración sin depender de mecanismos de señalización membranales especializados.[4]
En la mayoría de organismos en desarrollo, los sincítios raramente ocurren (como en el músculo esquelético), por lo que en general, los morfógenos son proteínas de señalización secretadas.
Algunos morfógenos
Véase también
- Biología del desarrollo
- Morfogénesis
- Factor de Transcripción
Referencias
- «Glosario | Biología, 7ma edición». www.curtisbiologia.com. Consultado el 21 de junio de 2021.
- Rojas, Mariana; Signore, Iskra A.; Mejías, Roberto (2014-03). «Morfógenos Durante el Desarrollo Embrionario de Vertebrados». International Journal of Morphology 32 (1): 319-326. ISSN 0717-9502. doi:10.4067/S0717-95022014000100051. Consultado el 21 de junio de 2021.
- «The chemical basis of morphogenesis». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences (en inglés) 237 (641): 37-72. 14 de agosto de 1952. ISSN 2054-0280. doi:10.1098/rstb.1952.0012. Consultado el 4 de mayo de 2020.
- Gilbert, Scott F., 1949-. Developmental biology (Eleventh edition edición). ISBN 978-1-60535-470-5. OCLC 945169933. Consultado el 4 de mayo de 2020.
Enlaces externos
| Control de autoridades |
|
|---|
 Datos: Q417084
Datos: Q417084