Raíz proteoide
En botánica, las raíces proteoides —a veces denominadas también raíces proteiformes o raíces en racimo—[1] son densos conglomerados de raíces laterales cortas y densamente espaciadas. Los conglomerados proteiformes se producen a intervalos de tiempo y distancias variables según la especie.[2] Pueden formar matas de dos hasta cinco centímetros de espesor, mejoran la absorción de nutrientes, posiblemente modificando químicamente el ambiente del suelo para incrementar la solubilización de los mismos. Como resultado, las plantas con raíces proteiformes pueden crecer en suelos con escasa cantidad de nutrientes, como aquellos con deficiencia de fósforo.[3] Fueron descritas por primera vez por Adolf Engler en 1894, luego que las descubriera en plantas de la familia de las proteáceas cultivadas en el Jardín Botánico de la Universidad de Leipzig. El término «raíz proteoide», no obstante, fue acuñado recién en 1960 por Helen Purnell tras descubrir que 44 especies pertenecientes a diez géneros de la mencionada familia presentaban este tipo de raíces.[4] Las raíces proteoides, si bien se descubrieron en las proteáceas, están ampliamente distribuidas taxonómicamente. De hecho, han sido descritas no solo en 27 géneros de esa familia, sino también en otras 30 especies de las familias betuláceas, casuarináceas, eleagnáceas, fabáceas, moráceas y mirtáceas. Estructuras similares a las raíces en racimo también se han descrito en las ciperáceas y restionáceas, pero su fisiología aún no está dilucidada.[2]

Muchas especies de plantas con raíces proteiformes tienen valor económico. Los cultivos con raíces proteiformes incluyen el lupino blanco (Lupinus albus),[5] utilizado como planta forrajera, varias especies de Banksia y Grevillea que se utilizan como plantas ornamentales y Macadamia integrifolia, cultivada por sus frutos comestibles.[6]
Morfología
Los conglomerados proteiformes pueden ser simples o compuestos. En el primer caso, se disponen aislados en series secuenciales y en el segundo caso, en forma de grupos de conglomerados. Estos grupos pueden ser cerrados, cuando de la raíz proteiforme nacen nuevos conglomerados proteiformes laterales; o abiertos, cuando los grupos se originan en raíces normales no proteiformes.[7] Ambos tipos de conglomerados pueden observarse en el mismo individuo, dependiendo de la edad del mismo. Así, en Gevuina avellana aparecen conglomerados proteiformes simples en la plántula y compuestos en los individuos adultos. En este último caso, pueden ser abiertos o cerrados, o de ambos tipos. Además, el patrón de ramificación de las raíces que portan conglomerados suele ser diferente según el tipo de suelo donde esté creciendo el árbol.[1]
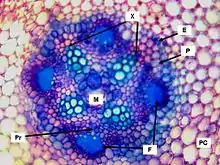
Las raicillas proteiformes presentan una morfología y anatomía más simple que las no proteiformes, además tienen un crecimiento determinado, comportándose como formas neoténicas, que detienen tempranamente su desarrollo primario, sin llegar a lignificarse.[8] A diferencia de las raíces convencionales, las raíces proteiformes carecen de caliptra, presentan crecimiento limitado y no forman tejidos secundarios. Al corte transversal presentan una rizodermis con abundantes pelos radicales, cuyo largo es igual a la longitud de la raicilla —alrededor de 1 cm— y con el extremo algo ramificado. Bajo ella hay una hilera de células con contenido más denso, la hipodermis, y luego una corteza primaria que presenta hasta cuatro estratos de grandes células parenquimáticas. La endodermis es uniestratificada y está formada por escasas células de gran tamaño que acumulan taninos en su interior. El periciclo, capa de células con actividad meristemática que rodea al cilindro vascular, está integrado por un solo estrato de células. El haz conductor radial es tetrarca, presenta cuatro radios de xilema con escasas células floemáticas intercaladas entre ellos. Hacia el extremo distal de la raicilla el haz conductor se reduce a diarca y luego desaparece.[2]
Debido a que las raicillas proteiformes no presentan crecimiento secundario en grosor, tienen una vida corta, que oscila entre los tres y cuatro meses. Cuando recién brotan no forman pelos radicales, sino que éstos aparecen más tarde, cuando las raicillas alcanzan su tamaño definitivo. En un comienzo son de color blanco y luego se tornan pardas, por la acumulación de taninos. Cuando están finalizando su ciclo de vida se forman estructuras con forma de clavo en su ápice, las cuales son muy frágiles y sobresalen de la superficie del conglomerado proteiforme. En estas estructuras normalmente no se forman pelos radicales, pero cuando aparecen, éstos son pequeños y deformes. Las estructuras claviformes señalan el inicio de la disgregación del conglomerado proteiforme, cuyas raicillas desaparecen totalmente, quedando solo la raíz proteiforme lignificada. Las estructuras claviformes progresivamente se convierten en un mucílago y son invadidas por una gran cantidad de microorganismos. Estas estructuras claviformes no se han descrito en las raíces proteoides de las especies de proteáceas investigadas tanto en Australia como en Sudáfrica, sino solamente en las especies estudiadas en América del Sur.[1]
Iniciación y crecimiento
Las raíces proteoides pueden representar desde el 40 hasta el 65 % de la biomasa total de la raíz,[9][10] y pueden contribuir hasta con el 80 % del crecimiento de la raíz de cada temporada cuando las plantas crecen en suelos pobres en nutrientes.[11][12] La iniciación del desarrollo de las raíces proteoides se halla bajo control genético, no obstante, la expresión de ese desarrollo está determinada por una cantidad de factores del ambiente.[13]

En las proteáceas que crecen en ambientes mediterráneos, el período de tiempo durante el cual se inician las raíces proteoides puede durar solo de dos a cuatro meses, en comparación con aquellas especies que habitan en regiones más húmedas donde las raíces proteoides funcionales pueden persistir mucho un intervalo mayor.[9][10] Sin embargo, las raíces en racimo puede estar fisiológicamente activas durante poco más de una semana en Lupinus albus,[14][15] y tal vez de dos a tres semanas en las especies investigadas de Hakea —por ejemplo, H. undulata y H. prostrata—[9][16][17] tras lo cual envejecen.
El color de la superficie de las raíces proteoides por lo general cambia de blanco a gris-marrón a medida que maduran, y ese cambio de color se ha utilizado para distinguir a las raicillas vivas de las senescentes, si bien las raíces marrones no siempre están muertas; de hecho, el eje subyacente de las raíces en racimo permanece intacto tras la senescencia de las raicillas.[18]
Desarrollo y relación con la disponibilidad de nutrientes
Cuando la humedad del suelo no restringe el crecimiento de la raíz, el desarrollo de las raíces proteoides se produce usualmente en las capas superiores del suelo, a continuación o incluso dentro de la capa de hojarasca que cubre la superficie del mismo.[9][19] El fósforo es un elemento clave que influye sustancialmente en la iniciación y el crecimiento de las raíces proteoides, por ejemplo, en la proteácea Macadamia integrifolia,[20] en la leguminosa Lupinus albus,[21] en la miricácea Myrica cerifera[18] y en la casuarinácea Gymnostoma papuanum.[22] Sin embargo, un bajo suministro de nitrógeno,[23] una limitación en el potasio,[24] o hierro,[25] también favorecen el desarrollo de las raíces proteoides.
La disponibilidad de fósforo se halla claramente implicada en el inicio y desarrollo de las raíces proteoides. Los racimos de raíces son más notorios cuando la oferta de fósforo en el suelo se halla restringida, y en la mayoría de las especies, la formación de este tipo de raíz disminuye si la disponibilidad de fósforo en el suelo aumenta.[9][26] De hecho, en Lupinus y en Myrica, el número de raíces proteoides se puede reducir mediante la aplicación de fósforo en las hojas, lo que demuestra que la concentración interna de fósforo, y no solo la oferta de este mineral en el suelo, puede influir en el inicio y desarrollo de estas estructuras. En Banksia ericifolia, la concentración de fósforo que resulta inhibitoria para la formación de raíces proteoides es más alta que la perjudicial para el crecimiento de plantas.[9] No obstante, en muchas especies las raíces en racimo se forman en las concentraciones de fósforo que comúnmente se encuentran en los suelos agrícolas (10 μm).[2] Se ha informado que la deficiencia de hierro también puede promover la formación de raíces proteoides en Lupinus consentinii, Ficus benjamina y Casuarina glauca pero no en Banksia ericifolia, L. albus o Alnus incana.[9][27]
Las condiciones que dan lugar a la formación de raíces proteoides varían ampliamente entre las especies de un mismo género. En Lupinus, ocho especies producen raíces proteoides, mientras que cuatro no lo hacen.[28] En Casuarina, cuatro especies producen raíces proteoides en respuesta a la deficiencia de fósforo, mientras que C. glauca los produce solo en respuesta a la deficiencia de hierro. Alnus rubra produce raíces proteoides en ausencia de hierro, mientras que A. incana los produce en presencia de fósforo y Alnus viridis no tiene la capacidad de producirlas bajo ningún tipo de condición.[29] Esta diversidad de respuestas, junto con el creciente número de familias en el que se han descubierto raíces proteoides, sugieren que las mismas se han originado independientemente en numerosas ocasiones durante la evolución de las plantas. Las características comunes en la morfología y en la fisiología de las mismas, sin embargo, indican que existe una combinación similar de las señales ambientales en la inducción de un racimo de raíces. Estas señales pueden ser provocadas por diferentes umbrales de nutrientes de la planta en las distintas especies.[2]
Función
La adquisición de fósforo por las raíces puede favorecerse a través del aumento de la superficie de sistema radicular o de la velocidad a la que el fósforo llega a la superficie de la raíz.[30] Una combinación de ambas estrategias permite a las especies que forman raíces proteoides adquirir más fósforo de los suelos que son deficientes en este nutriente, en comparación con las especies que no presentan raíces de racimo.[5][6] De hecho, las raíces proteoides adquieren diez veces más fósforo desde el suelo que las raíces que no poseen estas estructuras.[31] Aparentemente, los exudados producidos por este tipo de raíces mejoran la absorción de nutrientes a través de la modificación química del ambiente del suelo que incrementa la solubilización de los mismos.[6]
Las raíces de todas las plantas superiores son capaces de extraer nutrientes del suelo gracias a la exudación de compuestos químicos que hacen solubles a los minerales.[32][33][34] En particular, los ácidos tricarboxílicos —cítrico, málico, oxálico, entre otros— exudados por las raíces son importantes para promover la movilización del fósforo. En las especies con raíces proteoides la exudación de ácidos carboxílicos se incrementa en respuesta a la deficiencia de fósforo y, al mismo tiempo, decrece la tasa de varias rutas metabólicas que requieren fósforo como sustrato,[35] e incrementa la actividad de las enzimas involucradas en el suministro de compuestos carbonados y en la síntesis de ácidos carboxílicos.[36][37] Como resultado de la expresión conjunta de estos procesos, las plantas con raíces proteiformes pueden crecer en suelos con escasa cantidad de nutrientes, como aquellos con deficiencia de fósforo.[2][3][5][6]
Los estudios que utilizan suelos marcados con el isótopo 32P muestran que Lupinus albus puede extraer el fósforo ligado a las superficies de las arcillas que no se halla disponible para los cultivos que no forman raíces proteoides. Cuando se extrae y analiza la solución del suelo alrededor de las raíces proteoides, la misma presenta mayores niveles de fósforo, hierro y aluminio que la solución del resto del suelo. Además, la absorción de manganeso se incrementa con la formación de raíces proteoides como consecuencia de la exudación.[38][39] La solubilización de nutrientes también incrementa la disponibilidad de minerales para otras especies cuyas raíces pueden crecer entre las raíces proteoides. Por ejemplo, el trigo que se cultiva intercalado con L. albus es capaz de capturar dos veces más fósforo, nitrógeno y manganeso que cuando se cultiva en forma aislada.[6][40]
Véase también
- Morfología de la raíz (terminología)
- Raíz (botánica)
- Desarrollo de la raíz
- Meristema apical de la raíz
- Enraizamiento
Referencias
- Ramírez G.C., Valenzuela F.E. y San Martín P.C. 2004. Nuevos antecedentes sobre desarrollo temprano, morfología y anatomía de las raíces proteiformes de Gevuina avellana. Agro sur, vol.32, no.2, p.33-44
- Watt, Michelle and John R. Evans (1999). «Proteoid roots. Physiology and development». Plant Physiology 121: 317-323. doi:10.1104/pp.121.2.317. Consultado el 7 de noviembre de 2006.
- Grierson, P.F. and P. M. Attiwill (1989). «Chemical characteristics of the proteoid root mat of Banksia integrifolia L.». Australian Journal of Botany 37: 137-143. doi:10.1071/BT9890137.
- Purnell, Helen M. (1960). «Studies of the family Proteaceae: I. Anatomy and morphology of the roots of some Victorian species». Australian Journal of Botany 8 (1): 38-50. doi:10.1071/BT9600038.
- P.J. Hocking and S. Jeffery (2004). «Cluster-root production and organic anion exudation in a group of old-world lupins and a new-world lupin». Plant and Soil 258 (1): 135-150. doi:10.1023/B:PLSO.0000016544.18563.86.
- Shane, M.W. & Lambers, H. 2005. Cluster roots: A curiosity in context. Archivado el 20 de julio de 2017 en Wayback Machine. Plant Soil 274: 99-123.
- Lamont, B. 1983. Proteoid roots in the South African Proteaceae. South African Journal of Botany 49 (2): 103-123.
- Tahktajan, A. 1973. Evolution und Ausbreitung der Blütenpflanzen. Gustav Fischer Verlag, Stuttgart. 189 pp.
- Dinkelaker B., Hengeler C. & Marschner H. 1995. Distribution and function of proteoid root clusters and other root clusters. Bot. Acta 108, 183–200.
- Lamont B.B. 1981. Specialized roots of non-symbiotic origin in heathlands. In Ecosystems of the World, Vol. 9B, Heathlands and Related Shrublands. B. Analytical Studies. Ed. R L Specht. pp. 183–195. Elsevier Scientific, Amsterdam.
- Lamont B. 1972. ‘Proteoid’ roots in the legume Viminaria juncea. Search 3, 90-91.
- Lamont B. 1972. The morphology and anatomy of proteoid roots in the genus Hakea. Aust. J. Bot. 20, 155–174.
- Lambers H, Cramer M D, Shane M W, Wouterlood M, Poot P & Veneklaas E. 2003. Structure and functioning of cluster roots and plant responses to phosphate deficiency. Plant Soil 248, ix– xix.
- Hagstro J, James W M & Skene K.R. 2010. A comparison of structure, development and function in cluster roots of Lupinus albus L. under phosphate and iron stress. Plant Soil 232, 81–90.
- Keerthisinghe G, Hocking P, Ryan P. & Delhaize E. 1998. Proteoid roots of lupin (Lupinus albus L.): Effect of phosphorus supply on formation and spatial variation in citrate efflux and enzyme activity. Plant, Cell Environ. 21, 467– 478.
- Lamont B. 1976. The effects of seasonality and waterlogging on the root systems of a number of Hakea species. Aust. J. Bot. 24, 691–702.
- Shane M W, McCully M E & Lambers H. 2004. Tissue and cellular phosphorus storage during development of phosphorus toxicity in Hakea prostrata (Proteaceae). J. Exp. Bot. 55, 1033–1044. 2004b
- Louis I, Racette S & Torrey J.G. 1990. Occurrence of cluster roots in Myrica cerifera L. (Myricaceae) in water culture in relation to phosphorus nutrition. New Phytol. 115, 311–318.
- Jeffery D. W. 1967. Phosphate nutrition of Australian heath plants. I. The importance of proteoid roots in Banksia (Proteaceae). Aust. J. Bot. 15, 403–411.
- Aitken R L, Moody P W, Compton B L & Gallagher E.C. 1992. Plant and soil diagnostic tests for assessing the phosphorus status of seedling Macadamia integrifolia. Aust. J. Agr. Res. 43, 191–201.
- Gardner W K, Parbery D G & Barber D. A. 1982. The acquisition of phosphorus by Lupinus albus L. I. Some characteristics of the soil/root interface. Plant Soil 68, 19–32.
- Racette S, Louis I and Torrey J G 1990 Cluster roots formation by Gymnostoma papuanum (Casuarinaceae) in relation to aeration and mineral nutrient availability in water culture. Can. J. Bot. 68, 2564–2570.
- Lamont B. 1973. Factors affecting the distribution of proteoid roots within the root systems of two Hakea species. Aust. J. Bot. 21, 165–187.
- Liang R & Li C 2003 Differences in cluster-root formation and carboxylate exudation in Lupinus albus L. under different nutrient deficiencies. Plant Soil 248, 221–227.
- Zaıd E H, Arahou M, Diem H G & Morabet R. 2003. Is Fe deficiency rather than P deficiency the cause of cluster root formation in Casuarina species? Plant Soil 248, 229–235.
- Keerthisinghe G, Hocking PJ, Ryan PR, Delhaize E. 1998. Effect of phosphorus supply on the formation and function of proteoid roots of white lupin (Lupinus albus L.). Plant Cell Environ 21: 467–478).
- Arahou A, Diem HG (1997) Iron deficiency induces cluster (proteoid) root formation in Casuarina glauca. Plant Soil 196: 71–79
- Clements JC, White PF, Buirchell BJ (1993) The root morphology of Lupinus angustifolius in relation to other Lupinus species. Aust J Agric Res 44: 1367–1375
- Hurd T.M., Schwintzer C.R. 1996. Formation of cluster roots in Alnus incana ssp. rugosa and other Alnus species. Can J Bot 74:1684–1686
- Gardner W., Parbery D. & Barber D. 1982. [The acquisition of phosphorus by Lupinus albus L. II The effect of varying phosphorus supply and soil type on some characteristics of the soil/root interface. Plant Soil 68, 33-41.
- Jeschke D. & Pate J. 1995. Mineral nutrition and transport in xylem and phloem of Banksia prionotes (Proteaceae), a tree with dimorphic root morphology. J. Exp. Bot. 46, 895–905.
- Hinsinger P., Plassard C., Tang C. & Jaillard B. 2003. Origins of root mediated pH changes in the rhizosphere and their responses to environmental constraints: a review. Plant Soil 248, 43–59.
- Jones D., Dennis P. G., Owen A. G. & van Hees P. A. W. 2003. Organic acid behaviour in soils-misconceptions and knowledge gaps. Plant Soil 248, 31–41.
- Vance C. P., Uhde-Stone C. & Allen D. L. 2003. Phosphorus acquisition and use: critical adaptations by plants for securing a non-renewable source. New Phytol. 157, 423–447.
- Neumann G. & Romheld V. 1999. Root excretion of carboxylic acids and protons in phosphorus-deficient plants. Plant Soil 211, 121–130.
- Neumann G. & Romheld V. 2000. The release of root exudates as affected by the plant’s physiological status. In The Rhizosphere: Biochemistry and organic substances in the soil-plant interface. Eds. R. Pinton, Z. Varanini and P. Nannipieri. pp. 41– 93. Marcel Dekker, Inc., New York.
- Neumann G & Martinoia E. 2002. Cluster roots- an underground adaptation for survival in extreme environments. Trends Plant Sci. 7, 162–167.
- Braum S. & Helmke P. 1995. White lupin utilises soil phosphorus that is unavailable to soybean. Plant Soil 176, 95–100.
- Hocking P.J., Keerthisinghe G., Smith F.W., Randall P.J. 1998. A comparison of the ability of different crop species to access poorly-available soil phosphorus. In P Ando, K Fujita, T Mae, H Matsumoto, S Mori, J Sekiya, eds, Plant Nutrition for Sustainable Food Production and Environment. Kluwer Academic Publishers, Tokyo, pp 305–308
- Dinkelaker B., Hengeler C., Neumann G., Eltrop L., Marschner H. 1997. Root exudates and mobilization of nutrients. In H Rennenberg, W Eschrich, H Zeigler, eds, Trees: Contributions to Modern Tree Physiology. Backhuys, Leiden, The Netherlands, pp 441–451
Bibliografía
- Lambers, H. & Poot, P. (eds) 2003. Structure and Functioning of Cluster Roots and Plant Responses to Phosphate Deficiency. Kluwer Academic Publishers, Dordrecht.
- Shane, M.W. & Lambers, H. 2005. Cluster roots: A curiosity in context. Archivado el 20 de julio de 2017 en Wayback Machine. Plant Soil 274: 99-123.
- Lambers, H., Shane, M.W., Cramer, M.D., Pearse, S.J., & Veneklaas, E.J. 2006. Root structure and functioning for efficient acquisition of phosphorus: matching morphological and physiological traits. Ann. Bot. 98: 693-713.
| Control de autoridades |
|
|---|
 Datos: Q748536
Datos: Q748536