Red trófica
Una red trófica, (red alimentaria o un ciclo alimenticio) es la interconexión natural de las cadenas alimenticias y generalmente es una representación gráfica (usualmente una imagen) de quién se come a quién en una comunidad ecológica. Otro término para red alimenticia es un sistema de consumidor, recurso. Los ecólogos clasifican a los seres vivos de manera muy general en una de dos categorías llamadas niveles tróficos. Esta categorización comprende a:
- los autótrofos
- los heterótrofos.
Para mantener sus cuerpos, crecer, desarrollarse y reproducirse, los autótrofos producen materia orgánica desde sustancias inorgánicas, incluyendo minerales y gases como el dióxido de carbono. Esas reacciones químicas requieren energía, la cual principalmente proviene del sol, mayoritariamente de la fotosíntesis, aunque una cantidad puede provenir de aguas termales. Existe un gradiente entre los niveles tróficos que va desde los autótrofos estrictos que obtienen su única fuente de carbono de la atmósfera a los mixótrofos (como las plantas carnívoras) que son organismos autótrofos que pueden obtener materia orgánica parcialmente por otro método que no sea la atmósfera y hasta los heterótrofos estrictos que deben consumir a otros organismos para obtener materia orgánica. Las conexiones en una red alimenticia ilustran las rutas de consumo, en donde los heterótrofos obtienen materia orgánica al alimentarse de los autótrofos y otros heterótrofos. Una red alimenticia es, entonces, una ilustración simplificada de los varios métodos de alimentación que existen en un ecosistema y las conexiones que lo convierten en un sistema de intercambio único. Hay diferentes tipos de alimentación que se pueden dividir de forma general en herbívora, carnívora y parasitismo. Una parte de la materia orgánica consumida por los heterótrofos, como los azúcares, provee energía. Tanto los autótrofos como heterótrofos pueden ser de todos los tamaños, desde microscópicos hasta de toneladas de peso - desde cianobacteria hasta helechos gigantes, desde virus hasta ballenas azules.
Charles Elton fue el pionero en el concepto de los ciclos alimenticios, cadenas alimenticias y demás en su texto clásico de 1927, "Ecología Animal"; se reemplazó la cadena alimenticia de Elton por red alimenticia en textos subsecuentes. Elton organizó a las especies en grupos funcionales lo cual fue la base para la publicación clásica de Raymond Lindeman acerca de la dinámica trófica en 1942. Lindeman enfatizó la importancia de los descomponedores en el sistema de clasificación trófico. La noción de una red alimenticia tiene un origen histórico en los escritos de Charles Darwin y su terminología, incluyendo al "banco enredado", "red de la vida", "red de relaciones complejas" y "el continuo movimiento de las partículas de la tierra". Antes, en 1768, John Brucker describió a la naturaleza como una "red continua de vida".
Las redes alimenticias son representaciones "limitadas" de ecosistemas reales ya que agregan muchas especies en especies tróficas, que son grupos funcionales que tienen los mismos depredadores y presas en una red alimenticia. Los ecólogos usan esas simplificaciones en modelos cuantitativos o matemáticos de los sistemas consumidor-recurso para estudiar su dinámica.
Las redes alimenticias son mapas de los caminos descritos en el famoso banco enredado de Darwin y tienen una larga historia en la ecología. Al igual que los mapas de un lugar desconocido, las redes alimenticias parecen sorprendentemente complejas. A menudo se publicaban para demostrar justo eso. Sin embargo, estudios recientes muestran que las redes alimenticias de un amplio rango de comunidades terrestres, marinas y de aguas dulces comparten una importante lista de patrones.[1]: 669 |
Taxonomía de una red alimenticia

Las conexiones en una red alimenticia crean un mapa de las conexiones alimenticias (quién come a quién) en una comunidad ecológica. Ciclo alimenticio es el términdo anticuado sinónimo a red alimenticia. Los ecologistas agrupan a todas las formas de vida en dos grupos tróficos: los autótrofos y los heterótrofos. Los autótrofos obtienen energía (ya sea químicamente, capturando la energía del sol a través de la fotosíntesis o sin ella) que utilizan en la respiración. Los heterótrofos consumen biomasa en vez de producirla ya que metabolizan, crecen y añaden niveles de producción secundaria. Una red alimenticia ilustra un conjunto de consumidores polífagos heterotróficos que crean una red y circulan el flujo de energía y nutrientes desde una base productiva de autótrofos.[1][4][5]
El número de conexiones tróficas por consumidor es una medida de la conectancia de la red alimenticia. Las cadenas alimenticias son anidadas dentro de las conexiones tróficas de las redes alimenticias. Las cadenas alimenticias son rutas alimenticias lineales que siguen a consumidores monofagos desde la base hasta el máximo consumidor, que usualemente es un gran carnívoro.[6][7][8]
Las conexiones en una red alimenticia unen nodos, que son taxones biológicos llamados especies tróficas. Las especies tróficas son grupos funcionales que tienen los mismos depredadores y presas dentro de la red alimenticia. Ejemplos comunes de nodos incluyen parásitos, microbios, descomponedores, consumidores y depredadores, cada uno de los cuales contiene muchas posibles especies que pueden conectarse con otras especies tróficas.[9][10]
Niveles tróficos
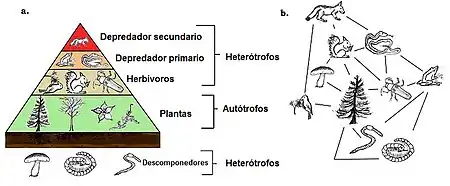
Las redes alimenticias tienen niveles y posiciones tróficas. Las especies basales, como las plantas, forman el primer nivel y son especies que no se alimentan de ninguna otra criatura en la red. Las especies basales pueden ser autótrofos o detritívoros, incluyendo "la descomposición de materia orgánica y sus microorganismos asociados que se definen como detrito, material micro inorgánico y microorganismos asociados así como material vegetal vascular."[12]: 94 La mayoría de los autótrofos capturan la energía solar a través de la clorofila, pero algunos otros (los quimiolitótrofos) la obtienen a través de la oxidación química de compuestos inórganicos y pueden crecer en ambientes oscuros, como la bacteria de azufre Thiobacillus, que vive en aguas termales sulfurosas. La cima tiene superpredadores que ninguna otra especie es capaz de matar directamente para utilizarlo como fuente de alimento. Los niveles intermedios están llenos de omnívoros que se alimentan de más de un nivel y causan que la energía fluya en muchas direcciones desde las especies basales[13]
En el esquema más simple, el primer nivel trófico son las plantas, luego los herbívoros y después los carnívoros en el tercer nivel. El nivel trófico es igual a uno más que la longitud de la cadena, que es el número de conexiones para llegar hasta la base. La base de la cadena tiene un número de cero.[1][14] Los ecologistas identifican relaciones de alimentación y organizan a las especies en categorías tróficas a través de un extenso análisis de contenido de vísceras de las diferentes especies. La técnica ha sido mejorada a través del uso de isótopos estables que mejoran el seguimiento del flujo de energía a través de la red.[15] Se pensaba que los organismos omnívoros eran raros, pero la evidencia sugiere lo contrario lo cual indica que las clasificaciones trópicas son más complejas.[16]
Dinámica trófica
El concepto de nivel trófico fue introducido en una publicación clave histórica de la dinámica trófica, en 1942 por Raymond Lindeman. La base de la dinámica trófica es la transferencia de energía de una parte del ecosistema a otra.[14][17] El concepto ha servido como herramienta cuantitativa, pero tiene varias limitaciones mayores incluyendo la precisión con la cual se puede relacionar a un organismo con un cierto nivel trófico. Los omnívoros, por ejemplo, no se restringen a un solo nivel. Sin embargo, investigaciones recientes revelan que los niveles tróficos sí existen, pero que "encima del nivel de los herbívoros, las redes alimenticias se clasifican mejor como una red compleja de omnívoros".[16]: 612
Una cuestión central en la literatura de la dinámica trófica es la naturaleza del control y regulación de los recursos y la producción. Los ecologistas utilizan modelos simplificados de una posición (productor, carnívoro, descomponedor). Usando esos modelos, los ecologistas han probado varios mecanismos de control ecológicos. Por ejemplo, los herbívoros normalmente tienen abundancia de recursos vegetales, lo que significa que el control de sus poblaciones se debe en gran parte a sus depredadores. Esto es conocido como la hipótesis de arriba-abajo o del mundo verde. De manera opuesta a la primera hipótesis, no toda la materia vegetal se puede consumir y la calidad nutricional y posibles defensas químicas de las plantas sugieren un modelo abajo-arriba.[18][19][20] Estudios recientes indican que ambos tipos de fuerzas pueden influenciar la estructura de una comunidad y que el grado con el que lo hacen es dependiente del contexto ambiental.[21][22] Esas interacciones complejas involucran a más de dos niveles tróficos.[23]
Otro ejemplo de interacción multitrófica es la cascada trófica, en la cual los depredadores ayudan a incrementar el crecimiento de las plantas al prevenir el consumo excesivo de las mismas por los herbívoros. Las conexiones en una red alimenticia ilustran relaciones tróficas directas entre especias, pero también hay efectos indirectos que pueden alterar la abundancia, distribución, o biomasa de los niveles tróficos. Por ejemplo, los depredadores que consumen herbívoros influencian indirectamente el control y regulación de la producción primaria en plantas. Aunque los depredadores no consuman las plantas de manera directa, regulan a la población de herbívoros que las consume. El efecto neto de las relaciones directas e indirectas se llama cascada trófica. Las cascadas tróficas se separan como cascadas por nivel de especies, donde solo un subconjunto de la dinámica de la red alimenticia es alterado por un cambio en número de población; así como las cascadas por nivel de comunidad donde un cambio en el número de población tiene un efecto dramático en toda la red.[24]
Flujo energético y biomasa
La ley de la conservación de la masa data del descubrimiento de Antoine Lavoisier de 1789 que enuncia que la masa no sea crea ni se destruye en las reacciones químicas. En otras palabras, la masa de cualquier elemento al principio de una reacción debe ser igual a la masa de ese mismo elemento al final de la reacción.[25]: 11 |
![]()
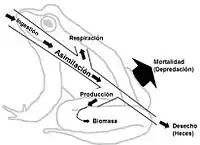
Derecha: Una cadena alimenticia de tres componentes, expandida (1. plantas, 2.herbívoros, 3.carnívoros) que ilustra la relación entre los diagramas de flujo alimenticios y la transformación energética. La energía se "degrada", dispersa y baja de calidad a lo largo de una cadena alimenticia. Abreviaturas: I=input (entrada), A=asimilación, R=respiración, NU=no utilizado, P=producción, B=biomasa.[26]
Las redes alimenticias ilustran el flujo de energía a través de conexiones tróficas. El flujo de energía es direccional, lo cual contrasta con el flujo de materia que es cíclico.[27] El flujo de energía "usualmente incluye producción, consumo, asimilación, perdidas de no asimilación (heces) y respiración (costo de mantenimiento)."[5]: 5 De modo muy general, el flujo de energía E se puede definir como la suma metabólica de la producción P y la respiración R de forma E=P+R.
La masa (o biomasa) de algo es igual a su contenido energético. La masa y la energía están relacionadas íntimamente. Sin embargo, la concentración y calidad de los nutrientes es variable. Muchas fibras de plantas, por ejemplo, no pueden ser digeridas por los herbívoros dejando a las comunidades de pastizales más limitadas en nutrientes que en otras donde las bacterias pudieran acceder a los nutrientes[28][29] "Los organismos usualmente extraen energía en la forma de carbohidratos, lípidos y proteínas. Esos polímeros tienen un rol doble tanto como fuentes de energía como bloques de construcción."[29]: 1230–1231 Las unidades en redes de flujo de energía son normalmente una medida de masa o energía por metro cuadrado por unidad de tiempo. Los diferentes consumidores tendrán diferentes eficiencias metabólicas en sus dietas. Cada nivel trófico transforma la energía en biomasa. Los diagramas de flujo de energía ilustran las tasas y eficiencia de transferencia de un nivel trófico a otro.[30][31]
Se da el caso de que la biomasa de cada nivel trófico disminuye de la base de la cadena hacia la cima. Esto es porque la energía se pierde en el ambiente en cada transferencia ya que aumente la entropía. Aproximadamente del 80 % al 90 % de la energía utilizada en los procesos vitales de los organismos se pierde como calor o desechos. Solo de 10 % a 20 % de la energía pasa al siguiente organismo.[32] la cantidad puede ser menor que 1 % en animales que consuman plantas poco digeribles y puede ser tan alta como 40 % en algunos casos de pláncton.[33] Las representaciones gráficas de la biomasa o productividad en cada nivel se llaman pirámide ecológicas o tróficas. La transferencia de energía de los productores primarios a los máximos consumidores puede ser caracterizada por diagramas de flujo de energía.[34]
Cadena alimenticia
Una medida utilizada comúnmente para cuantificar la estructura de las redes alimenticias es la longitud de la cadena alimenticia. Esta longitud es otra forma de describir a las redes alimenticias como una medida del número de especies encontradas como energía o nutrientes que se mueven de las plantas a los depredadores.[35]: 269 Hay diferentes formas de calcular esta longitud dependiendo de qué parámetros dinámicos sean considerados: conectancia, energía o interacción.[35] En su forma más simple, la longitud de una cadena es el número de conexiones entre un consumidor trófico y la base de la red. La longitud promedio de la red entera es el promedio de todas las longitudes de todas las cadenas en una red alimenticia.[4][13]
En un ejemplo simple de depredador-presa, un ciervo está alejado por un paso de las plantas que consume (longitud de cadena=1) y el lobo que se lo come está separado por dos pasos (longitud de cadena = 2). La influencia relativa que tienen los parámetros en las redes alimenticias se relacionan con interrogantes como:
Pirámides ecológicas

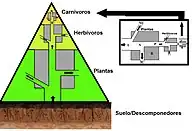

En una pirámide de números, el número de consumidores en cada nivel decrementa significativamente, de modo de que un solo superpredador (como un oso polar o un humano) es soportado por una cantidad mucho mayor de productores independientes entre sí. Usualmente hay un máximo de cuatro o cinco conexiones en una cadena alimenticia, aunque en sistemas acuáticos suele haber más. Eventualmente, toda la energía en una cadena termina convertida en calor.[4]
Las pirámides ecológicas ponen a los productores primarios en la base. Ilustran las diferentes propiedades de los ecosistemas, incluyendo el número de individuos por unidad de área, biomasa (g/m²), y energía (k cal m−2 año−1). El arreglo emergente de forma piramidal de los niveles tróficos con cantidades de energía que decremetan al removerse especies de la fuente de producción es uno de los patrones que se repite entre los ecosistemas planetarios.[3][1][38] El tamaño de cada nivel en la pirámide generalmente representa a la biomasa, que se puede medir como el peso seco de un organismo.[39] Los autótrofos tienen la mayor producción global de biomasa, con los microbios como contendiente que a veces los iguala o supera.[40][41]
La estructura piramidal puede variar a lo largo de los ecosistemas y el tiempo. En algunas instancias las pirámides de biomasa pueden invertirse. Este patrón se observa en ecosistemas acuáticos y arrecifes de corales. El patrón de la inversión de biomasa se atribuye a los diferentes tamaños de los productores. Las comunidades acuáticas son dominadas por prductores que son más pequeños y tienen muy altas tasas de crecimiento. Los productores acuáticos, como las algas planktónicas o acuáticas, carecen de la gran acumulación de crecimiento secundario que existe en los árboles maderosos de los ecosistemas terrestres. Sin embargo, son capaces de reproducirse lo suficientemente rápido para soportar una biomasa más grande. Esto invierte la pirámide. Los consumidores primarios tienen períodos de vida más grandes y crecimiento más lento que acumula más biomasa que los productores que consumen. Los fitoplánkton viven tan solo unos días, mientras que los zooplankton que consumen a los fitoplánkton viven por semanas y los peces que los consumen viven por varios años.[42] Además, la tasa de mortalidad de los depredadores acuáticos suele ser más baja que la de los consumidores pequeños, lo cual contribuye al patrón de pirámide invertida. La estructura poblacional, tasas de migración y refugios ambientales para las presas podrían ser otras causas de este patrón. Las pirámides de energía, sin embargo, siempre tendrían una forma de pirámide normal si se incluyen todas las fuentes de alimento y esto lo dicta la segunda ley de la termodinámica.[4][43]
Flujo de materia y reciclaje
Muchos de los elementos y minerales terrestres están contenidos dentro de los tejidos y dietas de los organismos. Por ende, los ciclos minerales y de nutrientes hacen un seguimiento de las rutas energéticas de las redes alimenticias. Los ecologistas emplean estequiometría para analizar los elementos principales encontados en todos los organismos: carbono (C), nitrógeno (N) y fósforo (P). Hay una gran diferencia entre muchos sistemas terrestres y acuáticos ya que las relaciones C:P y C:N ratios son mucho más altas en sistemas terrestres mientras que la relación N:P es igual en ambos sistemas.[44][45][46] Los nutrientes minerales son los recursos materiales que los organismos necesitan para el crecimiento, desarrollo y vitalidad. Las redes alimenticias describir las rutas de los ciclos de los minerales a manera en que estos fluyen a través de los organismos.[4][17] La mayoría de la producción primaria en un ecosistema no se consume, se recicla por desde el detrito a su forma de nutriente útil.[47] Muchos de los microorganismos del planeta están involucrados en la formación de minerales en un proceso llamado biomineralización.[48][49][50] Las bacterias que viven en sedimentos detríticos crean y circulan nutrientes y biominerales.[51] Aunque se hayan tratado a los modelos de red alimenticia y a los ciclos de nutrientes por separado, hay una fuerte correlación funcional entre los dos en términos de estabilidad, flujo, fuentes y reciclaje de nutrientes minerales.[52][53]
Tipos de redes alimenticias
Las redes alimenticias son una demostración diminuta de la complejidad de los ecosistemas reales. Por ejemplo, el número de especies en el planeta es cercano al orden de 107, con más del 95 % de estas especies siendo microbios e invertebrados, de los cuales relativamente pocos han sido clasificados por taxonomistas.[54][55][56] Se entiende explícitamente que los sistemas naturales son "resbalosos" y que las posiciones tróficas en una red alimenticia simplifican la complejidad de los sistemas reales que a veces están enfatizados en relaciones raras. La mayoría de los estudios se enfocan en las influencias más grandes donde sucede el grueso de la transferencia de energía.[18] "Estas omisiones y problemas son razones de preocupación, pero según las evidencias actuales, no representan dificultades irresolvibles."[1]: 669

Hay diferentes tipos o categorías de redes alimenticias:
- Red fuente - uno o más nodos, todos sus depredadores, toda la comida que estos consumen y así sucesivamente.
- Red hundida - uno o más nodos, todas sus presas, toda la comida que esas presas consumen y así sucesivamente.
- Red de comunidad o conectividad - un grupo de nodos y todas las conexiones de quién se come a quién.
- Red de flujo de energía - Flujos cuantificados de energía entre nodos a lo largo de conexiones entre un recurso y un consumidor.[1][39]
- Red paleoecológica - Una red que reconstruye ecosistemas desde el registro fósil.[57]
- Red funcional - Enfatiza la significancia funcional de ciertas conexiones que tengan mayor fuerza de interacción y por ende mayor peso en la organización de la comunidad, más aún que en los rutas de flujo de energía. Las redes funcionales tienen compartimientos, los cuales son subgrupos de la red más grande donde hay diferentes densidades y fuerzas de interacción.[37][58] Las redes funcionales enfatizan que "la importancia de mantener la integridad de una comunidad se refleja en su influencia en las tasas de crecimiento de otras poblaciones."[39]: 511
Dentro de esas categorías, las redes alimenticias se pueden organizan de acuerdo a los diferentes tipos de ecosistemas investigados. Por ejemplo, las redes alimenticias humanas, redes de agricultura, detríticas, de comida marina y acuática en general, del suelo, del ártico, terrestres y microbianas. Esas caracterizaciones surgen del concepto de ecosistema, lo cual asume que el fenómeno bajo investigación (interacciones y ciclos de retroalimentación) son suficientes para explicar patrones dentro de límites como la orilla del bosque, una isla, costa o alguna otra característica física pronunciada.[59][60][61]
Red detrítica
En una red detrítica, la materia animal y vegetal se fragmenta por los descomponedores, bacterias y hongos, y se mueve hacia los detritívoros y luego a los carnívoros.[62] A menudo hay relaciones entre la red detrítica y la red de "pasto". Los hongos producidos por descomponedores en la red detrítica se convierten una fuente de energía para ciervos, ardillas y hasta ratón. Los gusanos consumidos por aves son detritívoros al consumir hojas en descomposición.[63]
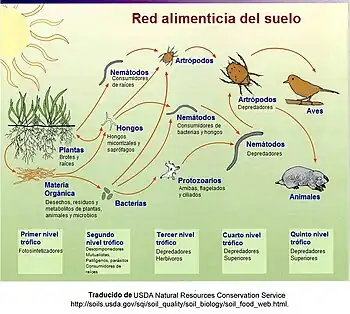
"El detrito puede ser definido de manera general como cualquier forma de materia orgánica no viva que incluye a diferentes tipos de tejido vegetal, (basura de hojas, madera muerta, algas, tejido animal (carroña), microbios muertos, heces, así como los productos secretados. La importancia relativa de esas formas de detrito varía en términos de origen, tamaño y composición a lo largo de diversos ecosistemas."[47]: 585
Redes alimenticias cuantitativas
Los ecólogos recolectan datos de los niveles tróficos y las redes alimenticias para modelar estadísticamente y calcular parámetros matemáticamente, como aquellos usados en otros tipos de análisis de redes para estudiar patrones y propiedades emergentes que estén compartidas entre ecosistemas. Hay diferentes dimensiones ecológicas que pueden ser mapeadas para crear redes alimenticias más complejas, incluyendo: composición de la especie (tipo de especie), la riqueza (número de especies), biomasa (el peso seco de plantas y animales), productividad (tasas de conversión de energía y nutrientes hacia crecimiento) y estabilidad (redes alimenticias en el tiempo). Un diagrama de red alimenticia que incluye composiciones de especies puede mostrar cómo el cambio en una sola especie puede afectar a muchas otras de manera directa e indirecta. Estudios de microcosmos se usan para simplificar la investigación en redes alimenticias para unidades semi-aisladas como troncos en descomposición y experimentos en laboratorio que utilizan organismos que se reproducen rápidamente, como dafnia alimentada por algas en ambientes controlados en recipientes de agua.[36][64]
Mientras que la complejidad de las conexiones reales en las redes alimenticias es complicada de descifrar, los ecólogos han encontrado que los modelos matemáticos son una herramienta invaluable para obtener información de la estructura, estabilidad y las leyes que rigen a los comportamientos y salidas de las redes alimenticias. "La teoría de redes alimenticias se centra alrededor de la idea de la conectancia".[65]: 1648 Las fórmulas cuantitativas simplifican la complejidad estructural de una red alimenticia. El número de conexiones o ligas tróficas (tL), por ejemplo, se convierte a un valor de conectancia:
- ,

donde, S(S-1)/2 es el máximo número de conexiones binarias entre S especies.[65] "La Conectancia (C) es la fracción de todas las posibles conexiones que pueden realizarse (L/S2) y representa una medida estándar para la complejidad de las redes alimenticias..."[66]: 12913 La distancia (d) entre cada par de especies es promediada para obtener la distancia media entre todos los nodos de una red (D)[66] y después se multiplica por el número total de conexiones (L) para obtener la densidad de conexiones (LD), la cual es influenciada por variables dependientes del tamaño, como la riquza de especies. Estas fórmulas son la base para comparar e investigar la naturaleza de los patrones no aleatorios en la estructura de las redes alimenticias en muchos tipos de eocsistemas[66][67]
Las leyes de escalamiento, complejidad, caos y correlaciones en patrón son atributos que comúnmente se utilizan para describir la complejidad de la estructura de una red alimenticia.[68][69]
La complejidad es una medida del número creciente de permutaciones y es también un término metafórico que expresa la intrazabilidad o los límites debido a la cantidad posible de posibilidades algorítimicas. En la terminología de redes alimenticias, la complejidad es producto el número de especies y su conectancia.[70][71][72] La conectancia es la "fracción de todas las posibles conexiones que se llevan a cabo en una red".[73]: 12917 Estos conceptos fueron derivados y estimulados a través del supuesto que la complejidad lleva a la estabilidad en las redes alimenticias, al incrementar el número de niveles tróficos en ecosistemas más ricos en especies. Esta hipótesis fue retada por modelos matemáticos que sugieren lo opuesto, pero más estudios han confirmado que la premisa se mantiene en sistemas reales.[70][74]
En diferentes niveles de la jerarquía de la vida, como en la estabilidad de las redes alimenticias, "la misma estructura general se mantiene aun cuando haya un flujo constante y cambio de componentes".[75]: 476 Entre más se aleje un sistema del equilibrio, más complejo se vuelve.[75] La complejidad tiene diversos significados en las ciencias de la vida y en la esfera pública por lo que se causa confusión en su aplicación como un término preciso con propósitos científicos analíticos.[72][76] La complejidad en las ciencias de la vida es definida por las "propiedades que emergen del conjunto de interacciones de comportamiento, biológicas, físicas y sociales, las cuales son afectadas, mantenidas o modificadas por los organismos vivos, incluyendo a los humanos".[77]: 1018
Varios conceptos han emergido del estudio de la complejidad en las redes alimenticias. La complejidad explica muchos principios acerca de la organización propia, no-linearidad, discontinuidad, emergencia y estabilidad en las redes alimenticias. La anidación, por ejemplo, se define como "un patrón de interacción en los cuales los especialistas interactúan con las especies que conforman subconjuntos perfectos con los cuales los generalistas interactúan",[78]: 575 "—esto es, la dieta de las especies más especializadas es un subconjunto de la siguiente más generalizada, y su dieta a la vez es un subconjunto de la que sigue, y así sigue."[79] Hasta recientemene, se pensaba que las redes alimenticias tenían poca estructura anidada, pero la evidencia empírica muestra que muchas redes ya publicadas tienen subredes anidadas en su composición[80] Como redes, exhiben propiedades matemáticas y estructurales usadas para describir otros tipos de sistemas complejos, como las redes de mundo pequeño y de libre escala. El atributo de mundo pequeño se refiere a muchos nodos conectados de manera leve, agrupamiento no aleatorio de nodos (como especies clave)[73][81] "Las redes ecológicas, especialmente aquellas de mutualismo, son generalmente muy heterogéneas, consistiendo de áreas con conexiones dispersas entre especies y áreas con conexiones muy concretas. Las regiones de alta densidad de conexiones pueden llamarse módulos, compartimientos o sub-grupos cohesivos... Dentro de las redes alimenticias, especialmente en sistemas marinos, la anidación parece relacionarse con el tamaño de los organismos porque las dietas de los organismos más pequeños tienden a anidarse en aquellas de los de mayor tamaño (Woodward & Warren 2007; YvonDurocher et al. 2008), y también se vuelven evidentes relaciones filogenéticas en donde los taxones relacionados se encuentran anidados debido a su historia evolutiva en común. (Cattin et al. 2004)."[82]: 257 "Los compartimientos en las redes evolutivas son subgrupos de taxones en los cuales pueden ocurrir muchas fuertes interacciones dentro de los subgrupos y pocas interacciones entre los subgrupos. Teóricamene, los compartimientos incrementan las estabilidad."[58]
Las redes alimenticias son complejas en la forma en que cambian de tamaño, tanto debido a las estaciones como geográficamente. Los componentes de las redes alimenticias, incluyendo a organismos y nutrientes minerales, cruzan los umbrales de los límites de ecosistemas. Esto ha llevado al concepto o área de estudio llamada subsidio de cruce de límites. .[59][60] "Esto lleva a anomalías, como que los cálculos de una red alimenticia arrojen que un ecosistema pueda soportar a una mitad de superpredadores carnívoros, sin especificar la dirección"[61] Sin embargo, se han identificado diferencias reales en estructura y función al compara diferentes tipos de redes alimenticias ecológicas.[83]
Historia de las redes alimenticias
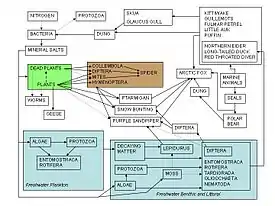
Las redes alimenticias les sirven a los ecologistas como marco para organizar las complejas redes de interacciones observadas entre las especies en la naturaleza alrededor del mundo. Una de las descripciones más tempranas de una cadena alimenticia fue la del académico medieval Al-Jahiz: "Todos los animales, en resumen, no pueden existir sin comida, ni tampoco puede el animal cazador escapar a ser cazado"[84]: 143 La descripción gráfica más temprana de una red alimenticia fue por Lorenzo Camerano en 1880, seguida independientemente por las de Pierce y su equipo en 1912 y la de Victor Shelford en 1913.[85][86] Dos redes alimenticias del arenque fueron descritas por Victor Summerhayes, Charles Elton[87] y Alister Hardy[88] en 1923 y 1924. Charles Elton subsecuentemente fue pionero en los conceptos de ciclos alimenticios, caenas alimenticias y tamaño alimenticio en su clásico libro de 1927 "Animal Ecology"; el 'ciclo alimenticio' descrito por Elton fue reemplazado por 'red alimenticia' en los textos ecológicos siguientes.[89] Después del uso de las redes alimenticias por Elton en 1927,[90] éstas se convirtieron en un concepto central en el campo de la ecología. Elton[89] ordenó a las especies en 'grupos funcionales' lo cual dio la base para el sistema de clasificación trófica en la publicación clásica de dinámica trófica de Raymond Lindeman en 1942.[17][37][91] La noción de una red alimenticia ha sido una base histórica en los escritos de Charles Darwin y su terminología, incluyendo un 'banco enredado', 'red de la vida', 'red de relaciones complejas' y en referencia a las descomposiciones por gusanos, Darwin hablaba del 'continuo movimiento de las partículas de la tierra'. Aún antes de esto, en 1768 John Bruckner describió a la naturaleza como una 'continuada red de vida'.[1][92][93][94]
Se ha incrementado el interés en las redes alimenticias después del estudio experimental y descriptivo de Robert Paine's de las orillas intermareales[95] sugiriendo que la complejidad de las redes alimenticias era clave para mantener la diversidad de las especies y la estabilidad ecológica. Muchos ecologistas teóricos, incluyendo a Sir Robert May[96] y Stuart Pimm[97] han sido llevados por este descubrimiento y otros a estudiar las propiedades matemáticas de las redes alimenticias.
Véase también
 Portal:Ecología. Contenido relacionado con Ecología.
Portal:Ecología. Contenido relacionado con Ecología. Portal:Ciencias de la Tierra. Contenido relacionado con Ciencias de la Tierra.
Portal:Ciencias de la Tierra. Contenido relacionado con Ciencias de la Tierra. Portal:Biología. Contenido relacionado con Biología.
Portal:Biología. Contenido relacionado con Biología.
Referencias
- Pimm, S. L.; Lawton, J. H.; Cohen, J. E. (1991). «Food web patterns and their consequences» (PDF). Nature 350 (6320): 669-674. doi:10.1038/350669a0. Archivado desde el original el 10 de junio de 2010.
- Kormondy, E. J. (1996). Concepts of ecology (4th edición). New Jersey: Prentice-Hall. p. 559. ISBN 0-13-478116-3.
- Proulx, S. R.; Promislow, D. E. L.; Phillips, P. C. (2005). «Network thinking in ecology and evolution». Trends in Ecology and Evolution 20 (6): 345-353. PMID 16701391. doi:10.1016/j.tree.2005.04.004. Archivado desde el original el 15 de agosto de 2011.
- Odum, E. P.; Barrett, G. W. (2005). Fundamentals of ecology. Brooks Cole. p. 598. ISBN 978-0-534-42066-6. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Benke, A. C. (2010). «Secondary production». Nature Education Knowledge 1 (8): 5.
- Allesina, S.; Alonso, D.; Pascual, M. «A general model for food web structure.». Science 320 (5876): 658-661. doi:10.1126/science.1156269. Archivado desde el original el 15 de mayo de 2016.
- Azam, F.; Fenche, T.; Field, J. G.; Gra, J. S.; Meyer-Reil, L. A.; Thingstad, F. (1983). «The ecological role of water-column microbes in the sea». Mar. Ecol. Prog. Ser. 10: 257-263. doi:10.3354/meps010257.
- Uroz, S.; Calvarus, C.; Turpault, M.; Frey-Klett, P. (2009). «Mineral weathering by bacteria: ecology, actors and mechanisms». Trends in Microbiology 17 (8): 378-387. PMID 19660952. doi:10.1016/j.tim.2009.05.004. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Williams, R. J.; Martinez, N. D. (2000). «Simple rules yield complex food webs.». Nature 404 (6774): 180-183. doi:10.1038/35004572. Archivado desde el original el 15 de marzo de 2012. Consultado el 5 de mayo de 2015.
- Post, D. M. (2002). «The long and short of food chain length». Trends in Ecology and Evolution 17 (6): 269-277. doi:10.1016/S0169-5347(02)02455-2. Archivado desde el original el 28 de julio de 2011.
- Odum, E. P.; Barrett, G. W. (2005). Fundamentals of Ecology (5th edición). Brooks/Cole, a part of Cengage Learning. ISBN 0-534-42066-4. Archivado desde el original el 20 de agosto de 2011. Consultado el 16 de junio de 2011.
- Tavares-Cromar, A. F.; Williams, D. D. (1996). «The importance of temporal resolution in food web analysis: Evidence from a detritus-based stream». Ecological Monographs 66 (1): 91-113. JSTOR 2963482.
- Pimm, S. L. (1979). «The structure of food webs». Theoretical population biology 16: 144-158. PMID 538731. doi:10.1016/0040-5809(79)90010-8. Archivado desde el original el 27 de septiembre de 2011.
- Cousins, S. «Ecologists build pyramids again.». New Scientist 1463: 50-54.
- McCann, K. (2007). «Protecting biostructure». Nature 446 (7131): 29. PMID 17330028. doi:10.1038/446029a. Archivado desde el original el 22 de julio de 2011.
- Thompson, R. M.; Hemberg, M.; Starzomski, B. M.; Shurin, J. B. (2007). «Trophic levels and trophic tangles: The prevalence of omnivory in real food webs.». Ecology 88 (3): 612-617. PMID 17503589. doi:10.1890/05-1454. Archivado desde el original el 15 de agosto de 2011.
- Lindeman, R. L. (1942). «The trophic-dynamic aspect of ecology». Ecology 23 (4): 399-417. doi:10.2307/1930126. Archivado desde el original el 29 de marzo de 2017. Consultado el 5 de mayo de 2015.
- Hairston, N. G.; Hairston, N. G. (1993). «Cause-effect relationships in energy flow, trophic structure, and interspecific interactions.». The American Naturalist 142 (3): 379-411. doi:10.1086/285546. Archivado desde el original el 20 de julio de 2011.
- Fretwell, S. D. (1987). «Food chain dynamics: The central theory of ecology?». Oikos 50: 291-301. doi:10.2307/3565489. Archivado desde el original el 28 de julio de 2011.
- Polis, G. A.; Strong, D. R. (1996). «Food web complexity and community dynamics.». The American Naturalist 147 (5): 813-846. doi:10.1086/285880. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Hoekman, D. «Turning up the head: Temperature influences the relative importance of top-down and bottom-up effects.». Ecology 91 (10): 2819-2825. doi:10.1890/10-0260.1.
- Schmitz, O. J. (2008). «Herbivory from individuals to ecosystems.». Annual Review of Ecology, Evolution and Systematics 39: 133-152. doi:10.1146/annurev.ecolsys.39.110707.173418.
- Tscharntke, T.; Hawkins, B., A., eds. (2002). Multitrophic Level Interactions. Cambridge: Cambridge University Press. p. 282. ISBN 0-521-79110-3.
- Polis, G. A. et al. (2000). «When is a trophic cascade a trophic cascade?». Trends in Ecology and Evolution 15 (11): 473-5. PMID 11050351. doi:10.1016/S0169-5347(00)01971-6.
- Sterner, R. W.; Small, G. E.; Hood, J. M. «The conservation of mass». Nature Education Knowledge 2 (1): 11.
- Odum, H. T. (1988). «Self-organization, transformity, and information». Science 242 (4882): 1132-1139. JSTOR 1702630. PMID 17799729. doi:10.1126/science.242.4882.1132.
- Odum, E. P. (1968). «Energy flow in ecosystems: A historical review». American Zoologist 8 (1): 11-18. doi:10.1093/icb/8.1.11.
- Mann, K. H. (1988). «Production and use of detritus in various freshwater, estuarine, and coastal marine ecosystems». Limnol. Oceanogr. 33 (2): 910-930. doi:10.4319/lo.1988.33.4_part_2.0910. Archivado desde el original el 25 de abril de 2012.
- Koijman, S. A. L. M.; Andersen, T.; Koo, B. W. (2004). «Dynamic energy budget representations of stoichiometric constraints on population dynamics». Ecology 85 (5): 1230-1243. doi:10.1890/02-0250.
- Anderson, K. H.; Beyer, J. E.; Lundberg, P. (2009). «Trophic and individual efficiencies of size-structured communities». Proc Biol Sci. 276 (1654): 109-114. PMC 2614255. PMID 18782750. doi:10.1098/rspb.2008.0951.
- Benke, A. C. (2011). «Secondary production, quantitative food webs, and trophic position». Nature Education Knowledge 2 (2): 2.
- Spellman, Frank R. (2008). The Science of Water: Concepts and Applications. CRC Press. p. 165. ISBN 978-1-4200-5544-3.
- Kent, Michael (2000). Advanced Biology. Oxford University Press US. p. 511. ISBN 978-0-19-914195-1.
- Kent, Michael (2000). Advanced Biology. Oxford University Press US. p. 510. ISBN 978-0-19-914195-1.
- Post, D. M. (1993). «The long and short of food-chain length». Trends in Ecology and Evolution 17 (6): 269-277. doi:10.1016/S0169-5347(02)02455-2.
- Worm, B.; Duffy, J.E. (2003). «Biodiversity, productivity and stability in real food webs». Trends in Ecology and Evolution 18 (12): 628-632. doi:10.1016/j.tree.2003.09.003.
- Paine, R. T. «Food webs: Linkage, interaction strength and community infrastructure». Journal of Animal Ecology 49 (3): 666-685. JSTOR 4220. doi:10.2307/4220.
- Raffaelli, D. (2002). «From Elton to mathematics and back again». Science 296 (5570): 1035-1037. PMID 12004106. doi:10.1126/science.1072080.
- Rickleffs, Robert, E. (1996). The Economy of Nature. University of Chicago Press. p. 678. ISBN 0-7167-3847-3.
- Whitman, W. B.; Coleman, D. C.; Wieb, W. J. (1998). «Prokaryotes: The unseen majority». Proc. Natl. Acad. Sci. USA 95 (12): 6578-83. PMC 33863. PMID 9618454. doi:10.1073/pnas.95.12.6578.
- Groombridge, B.; Jenkins, M. (2002). World Atlas of Biodiversity: Earth's Living Resources in the 21st Century. World Conservation Monitoring Centre, United Nations Environment Programme. ISBN 0-520-23668-8.
- Spellman, Frank R. (2008). The Science of Water: Concepts and Applications. CRC Press. p. 167. ISBN 978-1-4200-5544-3.
- Wang, H.; Morrison, W.; Singh, A.; Weiss, H. (2009). «Modeling inverted biomass pyramids and refuges in ecosystems». Ecological Modelling 220 (11): 1376-1382. doi:10.1016/j.ecolmodel.2009.03.005. Archivado desde el original el 7 de octubre de 2011.
- Pomeroy, L. R. «The strategy of mineral cycling». Annual Review of Ecology and Systematics 1: 171-190. JSTOR 2096770. doi:10.1146/annurev.es.01.110170.001131.
- Elser, J. J.; Fagan, W. F.; Donno, R. F.; Dobberfuhl, D. R.; Folarin, A.; Huberty, A.; et al. (2000). «Nutritional constraints in terrestrial and freshwater food webs». Nature 408 (6812): 578-580. doi:10.1038/35046058. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Koch, P. L.; Fox-Dobbs, K.; Newsom, S. D. «The isotopic ecology of fossil vertebrates and conservation paleobiology.». En Diet, G. P.; Flessa, K. W., eds. The Paleontological Society Papers 15. pp. 95-112.
- Moore, J. C.; Berlow, E. L.; Coleman, D. C.; de Ruiter, P. C.; Dong, Q.; Hastings, A.; et al. (2004). «Detritus, trophic dynamics and biodiversity». Ecology Letters 7 (7): 584-600. doi:10.1111/j.1461-0248.2004.00606.x.
- H. A., Lowenstam (1981). «Minerals formed by organisms.». Science 211 (4487): 1126-1131. JSTOR 1685216. PMID 7008198. doi:10.1126/science.7008198.
- Warren, L. A.; Kauffman, M. E. (2003). «Microbial geoengineers». Science 299 (5609): 1027-1029. JSTOR 3833546. PMID 12586932. doi:10.1126/science.1072076.
- Gonzalez-Acosta, B.; Bashan, Y.; Hernandez-Saavedra, N. Y.; Ascencio, F.; De la Cruz-Agüero, G. «Seasonal seawater temperature as the major determinant for populations of culturable bacteria in the sediments of an intact mangrove in an arid region.». FEMS Microbiology Ecology 55 (2): 311-321. doi:10.1111/j.1574-6941.2005.00019.x.
- DeAngelis, D. L.; Mulholland, P. J.; Palumbo, A. V.; Steinman, A. D.; Huston, M. A.; Elwood, J. W. (1989). «Nutrient dynamics and food-web stability.». Annual Review of Ecology and Systematics 20: 71-95. JSTOR 2097085. doi:10.1146/annurev.ecolsys.20.1.71.
- Twiss, M. R.; Campbell, P. G. C.; Auclair, J. (1996). «Regeneration, recycling, and trophic transfer of trace metals by microbial food-web organisms in the pelagic surface waters of Lake Erie.». Limnology and Oceanography 41 (7): 1425-1437. doi:10.4319/lo.1996.41.7.1425. Archivado desde el original el 25 de abril de 2012.
- May, R. M. (1988). «How many species are there on Earth?». Science 241 (4872): 1441-1449. PMID 17790039. doi:10.1126/science.241.4872.1441. Archivado desde el original el 11 de mayo de 2013. Consultado el 5 de mayo de 2015.
- Beattie, A.; Ehrlich, P. (2010). «The missing link in biodiversity conservation.». Science 328 (5976): 307-308. doi:10.1126/science.328.5976.307-c.
- Ehrlich, P. R.; Pringle, R. M. «Colloquium Paper: Where does biodiversity go from here? A grim business-as-usual forecast and a hopeful portfolio of partial solutions». Proceedings of the National Academy of Sciences 105 (S1): 11579-11586. PMC 2556413. PMID 18695214. doi:10.1073/pnas.0801911105.
- Dunne, J. A.; Williams, R. J.; Martinez, N. D.; Wood, R. A.; Erwin, D. H.; Dobson, Andrew P. (2008). «Compilation and Network Analyses of Cambrian Food Webs.». PLOS Biology 6 (4): e102. doi:10.1371/journal.pbio.0060102.
- Krause, A. E.; Frank, K. A.; Mason, D. M.; Ulanowicz, R. E.; Taylor, W. W. (2003). «Compartments revealed in food-web structure». Nature 426 (6964): 282-285. PMID 14628050. doi:10.1038/nature02115.
- Bormann, F. H.; Likens, G. E. (1967). «Nutrient cycling». Science 155 (3761): 424-429. doi:10.1126/science.155.3761.424. Archivado desde el original el 27 de septiembre de 2011.
- Polis, G. A.; Anderson, W. B.; Hold, R. D. (1997). «Toward an integration of landscape and food web ecology: The dynamics of spatially subsidized food webs». Annual Review of Ecology and Systematics 28: 289-316. doi:10.1146/annurev.ecolsys.28.1.289. Archivado desde el original el 2 de octubre de 2011.
- O'Neil, R. V. (2001). «Is it time to bury the ecosystem concept? (With full military honors, of course!)». Ecology 82 (12): 3275-3284. doi:10.1890/0012-9658(2001)082[3275:IITTBT]2.0.CO;2. Archivado desde el original el 25 de abril de 2012.
- Gönenç, I. Ethem; Koutitonsky, Vladimir G.; Rashleigh, Brenda (2007). Assessment of the Fate and Effects of Toxic Agents on Water Resources. Springer. p. 279. ISBN 978-1-4020-5527-0.
- Gil Nonato C. Santos; Alfonso C. Danac; Jorge P. Ocampo (2003). E-Biology II. Rex Book Store. p. 58. ISBN 978-971-23-3563-1.
- Elser, J.; Hayakawa, K.; Urabe, J. (2001). «Nutrient Limitation Reduces Food Quality for Zooplankton: Daphnia Response to Seston Phosphorus Enrichment.». Ecology 82 (3): 898-903. doi:10.1890/0012-9658(2001)082[0898:NLRFQF]2.0.CO;2.
- Paine, R. T. «Road maps of interactions or grist for theoretical development?». Ecology 69 (6): 1648-1654. doi:10.2307/1941141. Archivado desde el original el 28 de julio de 2011.
- Williams, R. J.; Berlow, E. L.; Dunne, J. A.; Barabási, A.; Martinez, N. D. (2002). «Two degrees of separation in complex food webs». Proceedings of the National Academy of Sciences 99 (20): 12913-12916. doi:10.1073/pnas.192448799.
- Banasek-Richter, C.; Bersier, L. L.; Cattin, M.; Baltensperger, R.; Gabriel, J.; Merz, Y.; et al. «Complexity in quantitative food webs». Ecology 90 (6): 1470-1477. doi:10.1890/08-2207.1. Archivado desde el original el 1 de junio de 2011.
- Briand, F.; Cohen, J. E. (1987). «Environmental correlates of food chain length.». Science (4829): 956-960. doi:10.1126/science.3672136. Archivado desde el original el 25 de abril de 2012.
- Neutel, A.; Heesterbeek, J. A. P.; de Ruiter, P. D. (2002). «Stability in real food webs: Weak link in long loops». Science 295 (550): 1120-1123. doi:10.1126/science.1068326. Archivado desde el original el 28 de septiembre de 2011.
- Leveque, C., ed. (2003). Ecology: From ecosystem to biosphere. Science Publishers. p. 490. ISBN 978-1-57808-294-0.
- Proctor, J. D.; Larson, B. M. H. (2005). «Ecology, complexity, and metaphor». BioScience 55 (12): 1065-1068. doi:10.1641/0006-3568(2005)055[1065:ECAM]2.0.CO;2. Archivado desde el original el 6 de octubre de 2011.
- Dunne, J. A.; Williams, R. J.; Martinez, N. D. (2002). «Food-web structure and network theory: The role of connectance and size». Proceedings of the National Academy of Sciences 99 (20): 12917-12922. doi:10.1073/pnas.192407699.
- Banašek-Richter, C.; Bersier, L.; Cattin, M.; Baltensperger, R.; Gabriel, J.; Merz, J.; et al. (2009). «Complexity in quantitative food webs». Ecology 90: 1470-1477. doi:10.1890/08-2207.1.
- Capra, F. (2007). «Complexity and life». Syst. Res. 24: 475-479. doi:10.1002/sres.848.
- Peters, R. H. (1988). «Some general problems for ecology illustrated by food web theory». Ecology 69 (6): 1673-1676. JSTOR 1941145.
- Michener, W. K.; Baerwald, T. J.; Firth, P.; Palmer, M. A.; Rosenberger, J. L.; Sandlin, E. A.; Zimmerman, H. (2001). «Defining and unraveling biocomplexity». BioScience 51: 1018-1023. doi:10.1641/0006-3568(2001)051[1018:daub]2.0.co;2. Archivado desde el original el 17 de agosto de 2011. Consultado el 5 de mayo de 2015.
- Bascompte, J.; Jordan, P. (2007). «Plant-animal mutualistic networks: The architecture of biodiversity.». Annu. Rev. Ecol. Evol. Syst. 38: 567-569. doi:10.1146/annurev.ecolsys.38.091206.095818. Archivado desde el original el 25 de octubre de 2009. Consultado el 3 de julio de 2011.
- Montoya, J. M.; Pimm, S. L.; Solé, R. V. (2006). «Ecological networks and their fragility». Nature 442 (7100): 259-264. doi:10.1038/nature04927. Archivado desde el original el 6 de julio de 2010. Consultado el 4 de julio de 2011.
- Michio, K.; Kato, S.; Sakato, Y. (2010). «Food webs are built up with nested subwebs». Ecology 91: 3123-3130. doi:10.1890/09-2219.1.
- Montoya, J. M.; Solé, R. V. (2002). «Small world patterns in food webs». Journal of Theoretical Biology 214 (3): 405-412. doi:10.1006/jtbi.2001.2460. Archivado desde el original el 5 de septiembre de 2011.
- Montoya, J. M.; Blüthgen, N; Brown, L.; Dormann, C. F.; Edwards, F.; Figueroa, D.; et al. (2009). «Ecological networks: beyond food webs». Journal of Animal Ecology 78: 253-269. doi:10.1111/j.1365-2656.2008.01460.x. Archivado desde el original el 16 de septiembre de 2011.
- Shurin, J. B.; Gruner, D. S.; Hillebrand, H. (2006). «All wet or dried up? Real differences between aquatic and terrestrial food webs». Proc. R. Soc. B 273 (1582): 1-9. PMC 1560001. PMID 16519227. doi:10.1098/rspb.2005.3377.
- Egerton, F. N. «A history of the ecological sciences, part 6: Arabic language science: Origins and zoological writings.». Bulletin of the Ecological Society of America 83 (2): 142-146.
- Egerton, FN (2007). «Understanding food chains and food webs, 1700-1970». Bulletin of the Ecological Society of America 88: 50-69. doi:10.1890/0012-9623(2007)88[50:UFCAFW]2.0.CO;2.
- Shelford, V. (1913). "Animal Communities in Temperate America as Illustrated in the Chicago Region". University of Chicago Press.
- Summerhayes, VS; Elton, CS (1923). «Contributions to the Ecology of Spitsbergen and Bear Island». Journal of Ecology 11: 214-286. doi:10.2307/2255864.
- Hardy, A. C. (1924). «The herring in relation to its animate environment. Part 1. The food and feeding habits of the herring with special reference to the east coast of England». Fisheries Investigation London Series II 7 (3): 1-53.
- Elton, C. S. (1927). Animal Ecology. London, UK.: Sidgwick and Jackson. ISBN 0-226-20639-4.
- Elton CS (1927) Animal Ecology. Republished 2001. University of Chicago Press.
- Allee, W. C. (1932). Animal life and social growth. Baltimore: The Williams & Wilkins Company and Associates.
- Stauffer, R. C. (1960). «Ecology in the long manuscript version of Darwin's "Origin of Species" and Linnaeus' "Oeconomy of Nature"». Proceedings of the American Philosophical Society 104 (2): 235-241. JSTOR 985662.
- Darwin, C. R. (1881). The formation of vegetable mould, through the action of worms, with observations on their habits.. London: John Murray.
- Worster, D. (1994). Nature's economy: A history of ecological ideas (2nd edición). Cambridge University Press. p. 423. ISBN 978-0-521-46834-3.
- Paine, RT (1966). «Food web complexity and species diversity». The American Naturalist 100: 65-75. doi:10.1086/282400.
- May RM (1973) Stability and Complexity in Model Ecosystems. Princeton University Press.
- Pimm SL (1982) Food Webs, Chapman & Hall.
Bibliografía
- Cohen, Joel E. (1978). Food webs and niche space. Monographs in Population Biology 11. Princeton, NJ: Princeton University Press. pp. xv+1-190. ISBN 978-0-691-08202-8.
- «Aquatic Food Webs». NOAA Education Resources. National Oceanic and Atmospheric Administration.
| Control de autoridades |
|
|---|
 Datos: Q1775153
Datos: Q1775153 Multimedia: Food chain / Q1775153
Multimedia: Food chain / Q1775153