Ribonucleasa H
La ribonucleasa H (RNasa H) es una familia de endonucleasas no específicas que catalizan la rotura de cadenas del ácido ribonucleico mediante un mecanismo hidrolítico. Está presente en prácticamente todos los organismos, desde bacterias y arqueas a eucariotas.
| Ribonucleasa H | ||||
|---|---|---|---|---|
 | ||||
| Estructuras disponibles | ||||
| PDB | ||||
| Identificadores | ||||
| Identificadores externos |
Bases de datos de enzimas
| |||
| Número EC | 3.1.26.4 | |||
| Número CAS | 9050-76-4 | |||
| Ortólogos | ||||
| Especies |
| |||
| PubMed (Búsqueda) |
| |||
| PMC (Búsqueda) |
| |||
La actividad ribonucleasa rompe los enlaces 3'-O-P del ácido ribonucleico hibridado con ADN para producir extremos 3'-hidroxil y 5'-fosfato. En la replicación del ADN, la RNasa H es responsable de la eliminación del cebador de ARN permitiendo terminar la síntesis completa del ADN
Estructura
La estructura tridimensional de la RNasa H consiste en cinco hojas beta antiparalelas rodeadas por hélices alfa.[1]
En algunas RNasas H, como la que se encuentra en el VIH-1, faltan una de las llamadas hélices C, hélice con carga positiva cuya forma protuberante aumenta su capacidad de unión a sustrato.[2] El sitio activo de este enzima está centrado en un dominio conservado DEDD (residuos D443, E478, D498 y D549) que lleva a cabo la hidrólisis del enlace fosfodiéster.[3] Normalmente se usa un catión magnesio como cofactor. Se especula con la necesidad de múltiples cationes para llevar a cabo este proceso.[4] La RNasa H también contiene una sitio de unión de ácidos nucleicos de 60 Å de longitud que interacciona con hasta 18 pares de bases del complejo ADN/ARN.
Función
Debido a que la RNasa H sólo degrada ARN hibridado con ADN, ha encontrado uso en biología molecular para hidrolizar y eliminar la plantilla de ARN usada en la síntesis de ADN complementario (ADNc) durante el proceso de transcripción inversa, así como en protocolos como ensayos de protección de nucleasa. También puede usarse para degradar ciertas cadenas de ARN en su unión a oligonucleótidos de ADNc, por ejemplo en la eliminación de la cola de poliadeninas del ARN mensajero hibridadas con oligo(dT), o la destrucción de ARN no codificante dentro o fuera de células vivas. La reacción se puede parar añadiendo un quelante de metales como el EDTA, el cual compleja y secuestra los iones metálicos como el Mg2+ en la mezcla de reacción.
Mecanismo
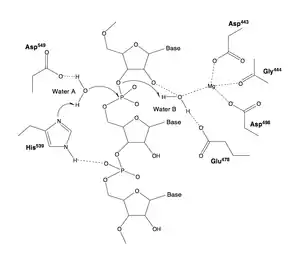
Genes
Los siguientes genes humanos codifican para proteínas con actividad RNasa H:
- ERVK6
- RNASEH1
- RNASEH2A
- RNASEH2B
- RNASEH2C
Rol en enfermedades
Se han asociado mutaciones en el gen RNASE2 con la enfermedad rara de origen genético conocida como síndrome de Aicardi-Goutières,[5] el cual se manifiesta a edades tempranas mediante síntomas neurológicos y dermatológicos.[6]
La actividad RNasa H de la transcriptasa inversa es una diana terapéutica importante, ya que es absolutamente necesaria para la proliferación de los retrovirus como el VIH-1 y el virus de la leucemia murina.[7][8] Inhibidores de este enzima podrían dar lugar a nuevos medicamentos contra enfermedades como el sida. Aunque no es una opción terapéutica efectiva, se ha visto que la 6'-desoxitioguanosina es capaz de inhibir la rotura de los enlaces fosfodiéster del ARN en los complejos ARN:ADN por parte de la RNasa H.[9]
VIH-1
La RNasa H existe como un dominio en el complejo heterodimérico de la transcriptasa inversa del VIH-1.[10] El VIH-1 tiene esta actividad enzimática necesaria para producir ADN de doble cadena a partir del ARN vírico. Durante la síntesis de este ADN, hay un paso inicial donde se forma un híbrido ADN:ARN como resultado de un intermedio de la replicación y por lo tanto dicho ARN debe ser eliminado por parte de la RNasa H antes de continuar con el proceso de retrotranscripción. La RNasa H tiene tres actividades: degradación no específica de la cadena positiva del ARN genómico, eliminación específica de la cadena negativa del cebador de ARNt y eliminación de la cadena positiva del cebador rico en polipurinas (CRP).[11] La RNasa H forma parte del proceso de cebado de la cadena positiva, pero no con el método de síntesis convencional de nuevos cebadores. En vez de esto, la RNasa H crea un cebador desde el CRP que es resistente a la degradación de la misma RNasa H. Al eliminar todas las bases excepto el CRP, éste es usado como un marcador del fina de la región U3 de su repetición terminal larga.[12]
Aplicaciones
Reacción en cadena de la polimerasa
La empresa IDT introdujo un método (PCR dependiente de RNasa H, o rhPCR) para incrementar la especificidad de la PCR usando RNasa H2, ya que ésta tiene poca actividad a temperatura ambiente y su máximo se encuentra en torno a 70 °C, para usarla como inhibidor de la PCR en reacciones de inicio en caliente. Los cebadores bloqueados contienen un único ribonucleótido.[13][14] La reacción de PCR sólo empieza cuando dicho ribonucleótido es degradado por la RNasa H2 a alta temperatura permitiendo la entrada de la polimerasa en el complejo.
Identificación de ácidos nucleicos
Se pueden identificar fragmentos de ADN mediante el reconocimiento y unión del ADN objetivo con una sonda de ARN. La RNasa H puede degradar selectivamente dichos complejos ARN:ADN, permitiendo la detección de dichas cadenas a nivel femtomolar.[15][16] Esta degradación se puede monitorizar mediante resonancia de plasmón de superficie para detectar la degradación de la sonda de microARN usada en la detección del virus de la influenza.
Referencias
- Schmitt TJ, Clark JE, Knotts TA (Diciembre de 2009). «Thermal and mechanical multistate folding of ribonuclease H». J Chem Phys 131 (23): 235101. PMID 20025349. doi:10.1063/1.3270167.
- Schultz SJ; Champoux JJ (June 2008). «RNase H activity: structure, specificity, and function in reverse transcription». Virus Res. 134 (1–2): 86-103. PMC 2464458. PMID 18261820. doi:10.1016/j.virusres.2007.12.007.
- Tadokoro T; Kanaya S (March 2009). «Ribonuclease H: molecular diversities, substrate binding domains, and catalytic mechanism of the prokaryotic enzymes». FEBS J. 276 (6): 1482-1493. PMID 19228197. doi:10.1111/j.1742-4658.2009.06907.x.
- Klumpp K; Hang JQ; Rajendran S et al. (December 2003). «Two-metal ion mechanism of RNA cleavage by HIV RNase H and mechanism-based design of selective HIV RNase H inhibitors». Nucleic Acids Res. 31 (23): 6852-9. PMC 290251. PMID 14627818. doi:10.1093/nar/gkg881.
- Crow, Y. J.; Leitch, A.; Hayward, B. E.; Garner, A.; Parmar, R.; Griffith, E.; Ali, M.; Semple, C.; Aicardi, J.; Babul-Hirji, R.; Baumann, C.; Baxter, P.; Bertini, E.; Chandler, K. E.; Chitayat, D.; Cau, D.; Déry, C.; Fazzi, E.; Goizet, C.; King, M. D.; Klepper, J.; Lacombe, D.; Lanzi, G.; Lyall, H.; Martínez-Frías, M. A. L.; Mathieu, M. L.; McKeown, C.; Monier, A.; Oade, Y.; Quarrell, O. W. (2006).
- Orcesi, S; La Piana, R; Fazzi, E (2009). «Aicardi-Goutieres syndrome». British Medical Bulletin 89: 183-201. PMID 19129251. doi:10.1093/bmb/ldn049.
- Mizuno M; Yasukawa K; Inouye K (February 2010). «Insight into the mechanism of the stabilization of moloney murine leukaemia virus reverse transcriptase by eliminating RNase H activity». Biosci. Biotechnol. Biochem. 74 (2): 440-2. PMID 20139597. doi:10.1271/bbb.90777.
- Coté ML; Roth MJ (June 2008). «Murine leukemia virus reverse transcriptase: structural comparison with HIV-1 reverse transcriptase». Virus Res. 134 (1–2): 186-202. PMC 2443788. PMID 18294720. doi:10.1016/j.virusres.2008.01.001.
- Krynetskaia NF; Krynetski EY; Evans WE (October 1999). «Human RNase H-mediated RNA cleavage from DNA-RNA duplexes is inhibited by 6-deoxythioguanosine incorporation into DNA». Mol. Pharmacol. 56 (4): 841-8. PMID 10496969.
- http://www.mdpi.com/1999-4915/2/4/900/pdf
- Klarmann GJ; Hawkins ME; Le Grice SF (2002). «Uncovering the complexities of retroviral ribonuclease H reveals its potential as a therapeutic target». AIDS Rev 4 (4): 183-94. PMID 12555693.
- Beilhartz GL; Götte M (March 2010). «HIV-1 Ribonuclease H: Structure, Catalytic Mechanism and Inhibitors». Viruses 2 (4): 900-926. doi:10.3390/v2040900.
- http://sg.idtdna.com/pages/products/genotyping/rnase-h2-dependent-pcr
- Dobosy JR; Rose SD; Beltz KR; Rupp SM; Powers KM; Behlke MA; Walder JA (2011). «RNase H-dependent PCR (rhPCR): improved specificity and single nucleotide polymorphism detection using blocked cleavable primers». BMC Biotechnol. 11: 80. PMID 21831278. doi:10.1186/1472-6750-11-80.
- Loo JF; Wang SS; Peng F; He JA; He L; Guo YC; Gu DY; Kwok HC; Wu SY; Ho HP; Xie WD; Shao YH; Kong SK (2015). «A non-PCR SPR platform using RNase H to detect MicroRNA 29a-3p from throat swabs of human subjects with influenza A virus H1N1 infection». Analyst 140: 4566-4575. PMID 26000345. doi:10.1039/C5AN00679A.
- Goodrich TT; Lee HJ; Corn RM (2004). «Direct detection of genomic DNA by enzymatically amplified SPR imaging measurements of RNA microarrays». J Am Chem Soc. 126: 4086-4087. PMID 15053580. doi:10.1021/ja039823p.