Sistema auditivo
El sistema auditivo es el conjunto de órganos que hacen posible el sentido del oído en un ser vivo. La función de nuestro sistema auditivo es, esencialmente, transformar las variaciones de presión originadas por la propagación de las ondas sonoras en el aire en impulsos eléctricos (variaciones de potencial), información que los nervios acústicos transmiten a nuestro cerebro para la asignación de significados.

La audición es el sentido que le permite a los órganos captar el sonido del ambiente. El sonido es el conjunto de ondas mecánicas de diversas amplitudes y frecuencias que se propagan por el aire. Los animales han desarrollado estructuras que permiten captar diferentes ondas sonoras. Por ejemplo, los mamíferos poseen oídos en la cabeza con tres compartimientos que transmiten las vibraciones sucesivamente hasta llegar a las células pilosas, que son sensibles a las vibraciones en una cierta frecuencia.
Podemos dividir el sistema auditivo en:
Proceso de la audición
El proceso de la audición implica que se conjuguen dos tipos de procesos:
- fisiológicos: Se capta todo sonido y lo transforman en impulsos eléctricos susceptibles de ser enviados al cerebro a través de los nervios auditivos. Los órganos que participan en esta parte del proceso conforman el sistema auditivo periférico.
- psicológicos: Interpretan estos sonidos, los reconocen y los dotan de significado. Los órganos que permiten esta percepción del sonido conforman el llamado sistema auditivo central.
Descripción del sistema
El oído externo canaliza las vibraciones del sonido hacia el tímpano, aumentando la presión sonora en la gama de frecuencias medias. El oído medio amplifica la presión de las vibraciones unas 20 veces. La base del estribo acopla las vibraciones en la cóclea a través de la ventana oval, que hace vibrar el líquido perilinfa (presente en todo el oído interno) y hace que la ventana redonda se abombe hacia fuera mientras la ventana oval se abomba hacia dentro.
El conducto vestibular y el conducto timpánico están llenos de perilinfa, y el conducto coclear más pequeño entre ambos está lleno de endolinfa, un fluido con una concentración de iones y un voltaje muy diferentes.[1][2][3] Las vibraciones perilinfáticas del conducto vestibular doblan las células externas del órgano de Corti (4 líneas) provocando la liberación de prestina en las puntas de las células. Esto hace que las células se alarguen y encojan químicamente (motor somático), y que los haces ciliados se desplacen, lo que, a su vez, afecta eléctricamente al movimiento de la membrana basilar (motor del haz ciliado). Estos motores (células ciliadas externas) amplifican las amplitudes de las ondas viajeras más de 40 veces.[4] Las células ciliadas externas (OHC) están mínimamente inervadas por el ganglio espiral en haces comunicativos recíprocos lentos (no mielinizados) (30+ pelos por fibra nerviosa); esto contrasta con las células ciliadas internas (IHC) que sólo tienen inervación aferente (30+ fibras nerviosas por un pelo) pero están fuertemente conectadas. Hay de tres a cuatro veces más OHC que IHC. La membrana basilar (BM) es una barrera entre las escamas, a lo largo de cuyo borde se sitúan las IHC y las OHC. La anchura y la rigidez de la membrana basilar varían para controlar las frecuencias mejor percibidas por las CCI. En la base coclear, la BM es más estrecha y rígida (frecuencias altas), mientras que en el ápice coclear es más ancha y menos rígida (frecuencias bajas). La membrana tectorial (TM) ayuda a facilitar la amplificación coclear estimulando la OHC (directamente) y la IHC (a través de las vibraciones de la endolinfa). La anchura y la rigidez de la TM son paralelas a las de la BM y, del mismo modo, contribuyen a la diferenciación de frecuencias.[5][6][7][8][9][10][11][12][13]
El complejo olivar superior (SOC), en el puente de Varolio, es la primera convergencia de los pulsos cocleares izquierdo y derecho. El SOC tiene 14 núcleos descritos; aquí se utilizan sus abreviaturas (véase Complejo olivar superior para sus nombres completos). MSO determina el ángulo de procedencia del sonido midiendo las diferencias de tiempo en la información izquierda y derecha. LSO normaliza los niveles de sonido entre los oídos; utiliza las intensidades del sonido para ayudar a determinar el ángulo del sonido. LSO inerva el IHC. Los VNTB inervan la OHC. Los MNTB inhiben el LSO a través de la glicina. Los LNTB son inmunes a la glicina y se utilizan para la señalización rápida. Los DPO son de alta frecuencia y tonotópicos. Los DLPO son de baja frecuencia y tonotópicos. Los VLPO tienen la misma función que los DPO, pero actúan en una zona diferente. PVO, CPO, RPO, VMPO, ALPO y SPON (inhibidos por la glicina) son diversos núcleos de señalización e inhibición.[14][15][16][17]
El cuerpo trapezoidal es donde la mayoría de las fibras del núcleo coclear (CN) se decusan (cruzan de izquierda a derecha y viceversa); este cruce ayuda a la localización del sonido.[18] El CN se divide en regiones ventrales (VCN) y dorsales (DCN). El VCN tiene tres núcleos. Las células tupidas transmiten información de sincronización, su forma promedia los temblores de sincronización. Las células estrelladas (chopper) codifican los espectros sonoros (picos y valles) mediante frecuencias de disparo neuronales espaciales basadas en la intensidad de la entrada auditiva (en lugar de la frecuencia). Las células pulposas tienen casi la mejor precisión temporal al disparar, decodifican el código temporal auditivo. El DCN tiene 2 núcleos. El DCN también recibe información del VCN. Las células fusiformes integran la información para determinar las claves espectrales de localización (por ejemplo, si un sonido procede de delante o de detrás). Cada una de las fibras nerviosas cocleares (más de 30 000) tienen una frecuencia más sensible y responden en una amplia gama de niveles.[19][20]
Simplificando, las señales de las fibras nerviosas son transportadas por las células arbusculares a las áreas binaurales del complejo olivar, mientras que los picos y valles de las señales son anotados por las células estrelladas, y la temporización de las señales es extraída por las células pulposas. El lemnisco lateral tiene tres núcleos: los núcleos dorsales responden mejor a la entrada bilateral y tienen respuestas de sintonía compleja; los núcleos intermedios tienen respuestas de sintonía amplia; y los núcleos ventrales tienen curvas de sintonía amplias y moderadamente complejas. Los núcleos ventrales del lemnisco lateral ayudan al colículo inferior (CI) a descodificar sonidos de amplitud modulada dando respuestas fásicas y tónicas (notas cortas y largas, respectivamente). El CI recibe entradas no mostradas, incluyendo áreas visuales (área pretectal: mueve los ojos ante el sonido. colículo superior: orientación y comportamiento hacia los objetos, así como movimientos oculares (sacada)), puente de Varolio (pedúnculo cerebeloso superior: conexión tálamo a cerebelo/escuchar sonido y aprender respuesta conductual), médula espinal (gris periacueductal: escuchar sonido y moverse instintivamente) y tálamo. Lo anterior es lo que implica al CI en la "respuesta de sobresalto" y los reflejos oculares. Más allá de la integración multisensorial, el CI responde a frecuencias específicas de modulación de la amplitud, lo que permite detectar el tono. El CI también determina las diferencias temporales en la audición binaural.[21] El núcleo geniculado medial se divide en ventral (células de retransmisión y células inhibidoras de retransmisión: frecuencia, intensidad e información binaural retransmitida topográficamente), dorsal (núcleos sintonizados amplios y complejos: conexión con la información somatosensorial) y medial (núcleos sintonizados amplios, complejos y estrechos: retransmisión de la intensidad y duración del sonido). El córtex auditivo (CA) permite percibir el sonido. El CA identifica los sonidos (reconocimiento del nombre del sonido) y también identifica el lugar de origen del sonido. El CA es un mapa topográfico de frecuencias con haces que reaccionan a diferentes armonías, tiempos y tonos. El AC del lado derecho es más sensible a la tonalidad, el AC del lado izquierdo es más sensible a las pequeñas diferencias secuenciales del sonido.[22][23] Las cortezas prefrontales rostromedial y ventrolateral están implicadas en la activación durante el espacio tonal y el almacenamiento de recuerdos a corto plazo, respectivamente.[24] La circunvolución de Heschl/giro temporal transversal incluye el área y la funcionalidad de Wernicke, está muy implicada en los procesos emoción-sonido, emoción-expresión facial y sonido-memoria. La corteza entorrinal es la parte del "sistema hipocampo" que ayuda y almacena los recuerdos visuales y auditivos.[25][26] El giro supramarginal (SMG) ayuda en la comprensión del lenguaje y es responsable de las respuestas compasivas. El SMG vincula los sonidos a las palabras con la circunvolución angular y ayuda en la elección de palabras. El SMG integra información táctil, visual y auditiva.[27][28]
Estructura

Oído externo
Los pliegues de cartílago que rodean el conducto auditivo externo se denominan pinna. Las ondas sonoras se reflejan y atenúan cuando chocan con el pabellón auditivo, y estos cambios proporcionan información adicional que ayudará al cerebro a determinar la dirección del sonido.
Las ondas sonoras entran en el conducto auditivo, un tubo aparentemente sencillo. El conducto auditivo amplifica los sonidos que están entre 3 y 12 kHz. El membrana timpánica, en el extremo más alejado del conducto auditivo externo marca el comienzo del oído medio.
Oído medio

Las ondas sonoras viajan a través del conducto auditivo externo y golpean la membrana timpánica o tímpano. Esta información de onda viaja a través de la cavidad del oído medio llena de aire a través de una serie de huesos delicados: el martillo, el yunque y el estribo. Estos huesecillos actúan como una palanca, convirtiendo las vibraciones sonoras del tímpano, de menor presión, en vibraciones sonoras de mayor presión en otra membrana más pequeña llamada ventana oval o ventana vestibular. El manubrium (mango) del martillo se articula con la membrana timpánica, mientras que la placa del pie (base) del estribo se articula con la ventana oval. La presión necesaria en la ventana oval es mayor que en la membrana timpánica, ya que el oído interno más allá de la ventana oval contiene líquido en lugar de aire. El reflejo de estapedio de los músculos del oído medio ayuda a proteger el oído interno de daños al reducir la transmisión de energía sonora cuando el músculo estapedio se activa en respuesta al sonido. El oído medio sigue conteniendo la información sonora en forma de onda; ésta se convierte en impulsos nerviosos en la cóclea.
Oído interno
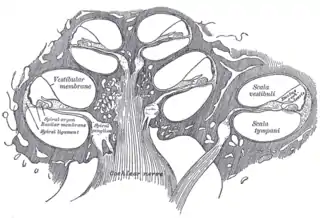
El oído interno está formado por la cóclea y varias estructuras no auditivas. La cóclea tiene tres secciones llenas de líquido (es decir, la scala media, scala tympani y scala vestibuli), y soporta una onda de líquido impulsada por la presión a través de la membrana basilar que separa dos de las secciones. Curiosamente, una sección, denominada conducto coclear o scala media, contiene endolinfa. En este conducto, situado en la membrana basilar, se encuentra el órgano de Corti, que transforma las ondas mecánicas en señales eléctricas en las neuronas. Las otras dos secciones se conocen como scala tympani y scala vestibuli. Están situadas dentro del laberinto óseo, que está lleno de un líquido llamado perilinfa, de composición similar al líquido cefalorraquídeo. La diferencia química entre los fluidos endolinfa y perilinfa es importante para la función del oído interno debido a las diferencias de potencial eléctrico entre los iones de potasio y calcio.[cita requerida]
La vista en planta de la cóclea humana (típica de todos los mamíferosios y de la mayoría de vertebradoss) muestra dónde se producen frecuencias específicas a lo largo de su longitud. La frecuencia es una función aproximadamente exponencial de la longitud de la cóclea dentro del órgano de Corti. En algunas especies, como los murciélagos y los delfines, la relación se amplía en zonas específicas para apoyar su capacidad de sonar activo.
Órgano de Corti

El órgano de Corti forma una cinta de epitelio sensorial que recorre longitudinalmente toda la scala media de la cóclea. Sus células ciliadas transforman las ondas de fluido en señales nerviosas. El viaje de innumerables nervios comienza con este primer paso; a partir de aquí, el procesamiento posterior conduce a una panoplia de reacciones y sensaciones auditivas.
Célula ciliada
Las células ciliadas son células columnares, cada una con un "haz ciliado" de 100-200 estereocilios especializados en la parte superior, de ahí su nombre. Existen dos tipos de células ciliadas específicas del sistema auditivo: las "células ciliadas internas" y las "células ciliadas externas". Las células ciliadas internas son los mecanorreceptores de la audición: transforman la vibración del sonido en actividad eléctrica en fibras nerviosas, que se transmite al cerebro. Las células ciliadas externas son una estructura motora. La energía sonora provoca cambios en la forma de estas células, lo que sirve para amplificar las vibraciones sonoras de una manera específica en función de la frecuencia. Sobre los cilios más largos de las células ciliadas internas descansa ligeramente la membrana tectorial, que se mueve hacia delante y hacia atrás con cada ciclo de sonido, inclinando los cilios, que es lo que provoca las respuestas eléctricas de las células ciliadas.
Las células ciliadas internas, al igual que las células fotorreceptoras del ojo, muestran un respuesta gradual, en lugar del picos típico de otras neuronas. Estos potenciales graduados no se rigen por las propiedades de "todo o nada" de un potencial de acción.
Llegados a este punto, cabe preguntarse cómo un meneo semejante de un haz de cilios desencadena una diferencia en el potencial de membrana. El modelo actual es que los cilios están unidos entre sí por "enlaces de puntas", estructuras que unen las puntas de un cilio a otro. Al estirarse y comprimirse, los enlaces de las puntas pueden abrir un canal iónico y producir el potencial receptor en la célula ciliada. Recientemente se ha demostrado que cadherina-23 CDH23 y protocadherina-15 PCDH15 son las moléculas de adhesión asociadas a estos enlaces de punta.[29] Se cree que un motor accionado por calcio provoca un acortamiento de estos enlaces para regenerar las tensiones. Esta regeneración de tensiones permite la aprehensión de estímulos auditivos prolongados.[30]
Neuronas
Las neuronas aferentes inervan las células ciliadas internas cocleares, en sinapsis donde el neurotransmisor glutamato comunica señales desde las células ciliadas a las dendritas de las neuronas auditivas primarias.
En la cóclea hay muchas menos células ciliadas internas que fibras nerviosas aferentes: muchas fibras nerviosas auditivas inervan cada célula ciliada. Las dendritas neuronales pertenecen a neuronas del nervio auditivo, que a su vez se une al nervio vestibular para formar el nervio vestibulococlear, o nervio craneal número VIII.[31]. La región de la membrana basilar que suministra las entradas a una fibra nerviosa aferente concreta puede considerarse su campo receptivo.
Las proyecciones eferentes del cerebro a la cóclea también desempeñan un papel en la percepción del sonido, aunque no se conoce bien. Las sinapsis eferentes se producen en las células ciliadas externas y en las dendritas aferentes (hacia el cerebro) de las células ciliadas internas.
Véase también
Referencias
- Tillotson, Joanne Kivela; McCann, Stephanie (2013). Flashcards de anatomía médica Kaplan. Kaplan Publishing. ISBN 978-1-60714-984-2.
- Ashwell, Ken (2016). Tarjetas didácticas de anatomía de Barron. Barron's Educational Series. ISBN 978-1-4380-7717-8.
- «¿Cómo funciona mi audición?». NZ Audiological Society. Archivado desde el original el 23 de agosto de 2019. Consultado el 27 de marzo de 2016.
- Zheng J, Shen W, He DZ, Long KB, Madison LD, Dallos P (Mayo 2000). «La prestina es la proteína motora de las células ciliadas externas de la cóclea». Nature 405 (6783): 149-55. Bibcode:..149Z 2000Natur.405 ..149Z. PMID 10821263. S2CID 4409772. doi:10.1038/35012009.
- Zwislocki JJ, Cefaratti LK (Noviembre 1989). «Membrana tectorial. II: Mediciones de rigidez in vivo». Hearing Research 42 (2-3): 211-27. PMID 2606804. S2CID 4706357. doi:10.1016/0378-5955(89)90146-9.
- Richter CP, Emadi G, Getnick G, Quesnel A, Dallos P (septiembre 2007). «Gradientes de rigidez de la membrana sectorial». Biophysical Journal 93 (6): 2265-76. Bibcode:...93.2265R 2007BpJ. ...93.2265R. PMC 1959565. PMID 17496047. doi:10.1529/biophysj.106.094474.
- Meaud J, Grosh K (Marzo 2010). «El efecto del acoplamiento longitudinal de la membrana tectorial y la membrana basilar en la mecánica coclear». The Journal of the Acoustical Society of America 127 (3): 1411-21. Bibcode:.127.1411M 2010ASAJ. .127.1411M. PMC 2856508. PMID 20329841. doi:10.1121/1.3290995.
- Gueta R, Barlam D, Shneck RZ, Rousso I (octubre 2006). «Medida de las propiedades mecánicas de la membrana tectorial aislada mediante microscopía de fuerza atómica». Proceedings of the National Academy of Sciences of the United States of America 103 (40): 14790-5. Bibcode:.10314790G 2006PNAS. .10314790G. PMC 1595430. PMID 17001011. doi:10.1073/pnas.0603429103.
- Freeman DM, Abnet CC, Hemmert W, Tsai BS, Weiss TF (Junio 2003). «Propiedades dinámicas del material de la membrana tectorial: resumen». Hearing Research 180 (1-2): 1-10. PMID 12782348. S2CID 24187159. doi:10.1016/S0378-5955(03)00073-X.
- Legan PK, Lukashkina VA, Goodyear RJ, Kössi M, Russell IJ, Richardson GP (Octubre 2000). «Una deleción dirigida en la alfa-tectorina revela que la membrana tectorial es necesaria para la ganancia y el tiempo de la retroalimentación coclear». Neuron 28 (1): 273-85. PMID 11087000. doi:10.1016/S0896-6273(00)00102-1.
- Canlon B (1988). «El efecto del trauma acústico sobre la membrana tectorial, los estereocilios y la sensibilidad auditiva: posibles mecanismos subyacentes al daño, la recuperación y la protección». Scandinavian Audiology. Supplementum 27: 1-45. PMID 3043645.
- Zwislocki JJ (1979). «Membrana tectorial: un posible efecto de agudización en el análisis de frecuencias en la cóclea». Acta Oto-Laryngologica 87 (3-4): 267-9. PMID 443008. doi:10.3109/00016487909126419.
- Teudt IU, Richter CP (October 2014). «Rigidez de la membrana basilar y la membrana tectorial en el ratón CBA/CaJ». Journal of the Association for Research in Otolaryngology 15 (5): 675-94. PMC 4164692. PMID 24865766. doi:10.1007/s10162-014-0463-y.
- Thompson AM, Schofield BR (Noviembre 2000). «Proyecciones diferentes del complejo olivar superior». Microscopy Research and Technique 51 (4): 330-54. PMID 11071718. S2CID 27179535.
- Oliver DL (Noviembre 2000). «Proyecciones eferentes ascendentes del complejo olivar superior». Microscopy Research and Technique 51 (4): 355-63. PMID 11071719. S2CID 36633546.
- Moore JK (Noviembre 2000). «Organización del complejo olivar superior humano». Microscopy Research and Technique 51 (4): 403-12. PMID 11071722. S2CID 10151612. doi:10.1002/1097-0029(20001115)51:4<403::AID-JEMT8>3.0.CO;2-Q.
- Yang L, Monsivais P, Rubel EW (Marzo 1999). «El núcleo olivar superior y su influencia en el núcleo laminar: una fuente de retroalimentación inhibitoria para la detección de coincidencias en el tronco encefálico auditivo aviar». The Journal of Neuroscience 19 (6): 2313-25. PMC 6782562. PMID 10066281. doi:10.1523/JNEUROSCI.19-06-02313.1999.
- Paolini AG, FitzGerald JV, Burkitt AN, Clark GM (September 2001). «Temporal processing from the auditory nerve to the medial nucleus of the trapezoid body in the rat». Hearing Research 159 (1-2): 101-16. PMID 11520638. S2CID 25279502. doi:10.1016/S0378-5955(01)00327-6.
- Bajo VM, Merchán MA, Malmierca MS, Nodal FR, Bjaalie JG (Mayo 1999). «Organización topográfica del núcleo dorsal del lemnisco lateral en el gato». The Journal of Comparative Neurology 407 (3): 349-66. PMID 10320216. S2CID 25724084.
- Young ED, Davis KA (2002). «Circuitería y función del núcleo coclear dorsal». En Oertel D, Fay RR, Popper AN, ed. Funciones integradoras en la vía auditiva de los mamíferos. Springer Handbook of Auditory Research 15. Nueva York, NY: Springer. pp. 160-206. ISBN 978-1-4757-3654-0. doi:10.1007/978-1-4757-3654-0_5.
- Oliver DL (2005). «Organización neuronal en el colículo inferior». En Winer JA, Schreiner CE, ed. El colículo inferior. New York, NY: Springer. pp. 69-114. ISBN 978-0-387-27083-8. doi:10.1007/0-387-27083-3_2.
- Janata P, Birk JL, Van Horn JD, Leman M, Tillmann B, Bharucha JJ (Diciembre 2002). «La topografía cortical de las estructuras tonales subyacentes a la música occidental». Science 298 (5601): 2167-70. Bibcode:..298.2167J 2002Sci. ..298.2167J. PMID 12481131. S2CID 3031759. doi:10.1126/science.1076262.
- Morosan P, Rademacher J, Schleicher A, Amunts K, Schormann T, Zilles K (Abril 2001). «Corteza auditiva primaria humana: subdivisiones citoarquitectónicas y mapeo en un sistema de referencia espacial». NeuroImage 13 (4): 684-701. PMID 11305897. S2CID 16472551. doi:10.1006/nimg.2000.0715.
- Romanski LM, Tian B, Fritz J, Mishkin M, Goldman-Rakic PS, Rauschecker JP (Diciembre 1999). «Dual streams of auditory afferents target multiple domains in the primate prefrontal cortex». Nature Neuroscience 2 (12): 1131-6. PMC 2778291. PMID 10570492. doi:10.1038/16056.
- Badre D, Wagner AD (octubre 2007). «Corteza prefrontal ventrolateral izquierda y el control cognitivo de la memoria». Neuropsychologia 45 (13): 2883-901. PMID 17675110. S2CID 16062085. doi:10.1016/j.neuropsychologia.2007.06.015.
- Amunts K, Kedo O, Kindler M, Pieperhoff P, Mohlberg H, Shah NJ, Habel U, Schneider F, Zilles K (Diciembre 2005). «Mapeo citoarquitectónico de la amígdala humana, región del hipocampo y corteza entorrinal: variabilidad intersujeto y mapas de probabilidad». Anatomy and Embryology 210 (5-6): 343-52. PMID 16208455. S2CID 6984617. doi:10.1007/s00429-005-0025-5.
- Penniello MJ, Lambert J, Eustache F, Petit-Taboué MC, Barré L, Viader F, Morin P, Lechevalier B, Baron JC (Junio 1995). «Un estudio PET de la neuroanatomía funcional del deterioro de la escritura en la enfermedad de Alzheimer. The role of the left supramarginal and left angular gyri». Brain: A Journal of Neurology. 118 ( Pt 3) (3): 697-706. PMID 7600087. doi:10.1093/brain/118.3.697.
- Stoeckel C, Gough PM, Watkins KE, Devlin JT (October 2009). «Participación del giro supramarginal en el reconocimiento visual de palabras». Cortex; A Journal Devoted to the Study of the Nervous System and Behavior 45 (9): 1091-6. PMC 2726132. PMID 19232583. doi:10.1016/j.cortex.2008.12.004.
- Lelli A, Kazmierczak P, Kawashima Y, Müller U, Holt JR (Agosto 2010). «El desarrollo y la regeneración de la transducción sensorial en las células ciliadas auditivas requiere la interacción funcional entre cadherina-23 y protocadherina-15». The Journal of Neuroscience 30 (34): 11259-69. PMC 2949085. PMID 20739546. doi:10.1523/JNEUROSCI.1949-10.2010.
- Peng AW, Salles FT, Pan B, Ricci AJ (Noviembre 2011). «Integrando los mecanismos biofísicos y moleculares de la mecanotransducción de las células ciliadas auditivas». Nature Communications 2: 523. Bibcode:..2..523P 2011NatCo. ..2..523P. PMC 3418221. PMID 22045002. doi:10.1038/ncomms1533.
- Meddean - CN VIII. Nervio vestibulococlear
| Control de autoridades |
|
|---|
 Datos: Q821413
Datos: Q821413 Multimedia: Auditory system / Q821413
Multimedia: Auditory system / Q821413