Sistema visual
El sistema visual es la parte del sistema nervioso central que les brinda a los organismos la habilidad de procesar detalle visual, así como habilitar la formación de varias funciones de respuesta de foto sin imagen. Detecta e interpreta información de la luz visible para crear una representación del ambiente alrededor. El sistema visual lleva a cabo una serie de tareas complejas, incluyendo la recepción de luz y la formación de representaciones monoculares; la acumulación de una percepción binocular nuclear de un par de dos proyecciones dimensionales; la identificación y categorización de objetos visuales; evaluando distancias hacia y entre objetos; y guiando los movimientos del cuerpo en relación con el objeto visto. El proceso psicológico de información visual se conoce como percepción visual, una falta de la cual se llama ceguera. Las funciones visuales sin información de imágenes, independiente de la percepción visual, incluye el reflejo de luz pupilar (RLP) y el fotoentrenamiento circadiano.
Más que nada, este artículo describe el sistema visual de mamíferos, en especial en humanos, pero también en animales “mayores” que tienen un sistema visual similar.
Resumen del sistema
Mecánica
La córnea y los lentes del cristalino refractan la luz en una pequeña imagen y brilla sobre la retina. La retina traduce esta imagen en un impulso eléctrico usando los bastones y los conos. Los nervios ópticos son los que cargan estos impulsos a través del canal óptico. Cuando llega al quiasma óptico las fibras nerviosas se cruzan (izquierda se hace derecha). Después las fibras se ramifican y terminan en tres lugares.[1][2][3][4][5][6][7]
Neuronal

La mayoría terminan en el núcleo lateral geniculado (LGN). Previo a que el LGN llegue a los pulsos del V1 en el córtex visual (primario) mide el rango de objetos y etiqueta a cada objeto mayor con una etiqueta de velocidad. Estas etiquetas predicen el movimiento de los objetos.
El LGN también manda información (carácter) al V2 y V3.[8][9][10][11][12]
El V1 trabaja la detección de los bordes para entender la organización de este espacio (inicia en 40 milisegundos, enfocando solo en pequeños espacios y cambios de color. Luego, 100 milisegundos después, al recibir el LGN traducido, con la información del V2 y V3, también comienza enfocando a organización global).
El V2 se mueve adelante (directamente a la vía pulvinar) impulsa al V1 y los recibe.
El pulvinar es el responsable de la atención visual. El V2 tiene casi la misma función que el V1, sin embargo, también se encarga de los contornos ilusorios, determinando la profundidad comparando los pulsos derechos e izquierdos (imágenes 2D), distinguiendo el primer plano. V2 se conecta a V1-V5.
El V3 ayuda a procesar el “movimiento global” (dirección y velocidad)de los objetos. El V3 se conecta con el V1 (tenuemente), V2, y el córtex temporal inferior.[13][14]
El V4 reconoce las figuras simples, se conecta firmemente con el V1, V2, V3, LGN y el pulvinar.[15] salidas del V5 incluyen al V4 y el área alrededor, y el movimiento motor de las cortezas oculares (el área intraparietal del ojo).
Las funciones del V5 son similares a la de los otros V´s, sin embargo, integra el movimiento de los objetos locales a un movimiento local global de un nivel complejo. V6 trabaja en la conjunción con el V5 y un análisis al movimiento. El V5 analiza el auto-movimiento, donde el V6 analiza el movimiento de los objetos relativos a un segundo plano. El input primario del V6 es V1, con las aportaciones del V5. El V6 atrapa el mapa topográfico de la visión. El V6 se desconecta de la región que está alrededor de este (V6A). V6A tiene conexiones al movimiento de los brazos vórtices, incluyendo el córtex premotor.[16][17]
El giro temporal inferior reconoce figuras complejas, objetos y caras, o en conjunto con el hipocampos crea nuevas memorias.[18] El pretectum o zona pretectal está compuesta por siete núcleos únicos. Núcleos pretectales anteriores, posteriores y medios inhiben el dolor (indirectamente), ayudan en el REM y ayudan a acomodar reflejos, respectivamente.[19] El núcleo Edinger-Westphal modera la dilatación de la pupila y ayuda (pues provee fibras parasimpáticas) en la convergencia de los ojos y el ajuste de los lentes.[20] Los núcleos de la cintilla óptica se involucran en un suave búsqueda de movimiento ocular y de acomodar el reflejo, al igual que REM.
El núcleo supraquiasmático es la región del hipotálamo que detiene la producción de melatonina (indirectamente) a primera vista.[21]
Estructura
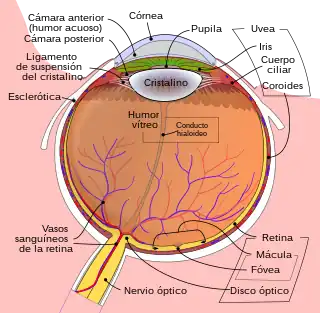
- El ojo, especialmente la retina.
- El nervio óptico.
- El quiasma óptico.
- El tracto óptico.
- El cuerpo geniculado lateral.
- El córtex visual.
- El córtex de asociación visual.
Estos se dividen en vías anteriores y posteriores. La vía anterior visual se refiere a las estructuras involucradas en la visión antes del núcleo geniculado lateral. La vía posterior visual se refiere a las estructuras después de este punto.
Ojo
La luz que entra al ojo es refractada a medida que pasa a través de la córnea. Luego pasa a través de la pupila (controlada por el iris) y se refracta aún más por la lente. La córnea y lente actúan juntos como una lente compuesta para proyectar una imagen invertida en la retina.
Retina
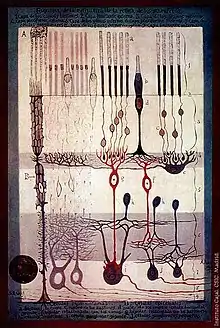
La retina consiste de un largo número de células fotorreceptoras cuales contienen moléculas de proteínas llamadas opsinas. En los humanos, hay dos tipos de opsinas que están involucradas con la visión, bastones de opsinas y opsinas coníferas. (Hay un tercer tipo, melanopsina, que está en las células ganglionares de la retina (RGC),parte del reloj biológico mecánico, probablemente no está envuelto con la visión consciente, las RGC se proyecta al núcleo olivar pretectal (ON)[22]). Una opsina absorbe un fotón (partícula de luz) y transmite señal a una célula a través de una señal de camino traductora, llegando en la hiperpolarización de un fotoreceptor.
Los bastones y los conos difieren en su función. Los bastones se encuentran principalmente en la periferia de la retina y son utilizados para ver en niveles bajos de luz. Los conos se encuentran principalmente en el centro (o fóvea) de la retina.[23] Existen tres tipos de conos que difieren en las longitudes de onda de la luz que absorben; normalmente se les llama corto o azul, mediano o verde y largo o rojo. Los conos se utilizan principalmente para distinguir el color y otras características del mundo visual en niveles normales de luz.[23]
En la retina, los fotorreceptores hacen sinapsis directo sobre las células bipolares, las cuales dirigen la sinapsis hacia las células ganglionares de la capa más exterior, la cual conduce el potencial de acción al cerebro. Una parte significativa del proceso visual nace de los patrones de comunicación entre neuronas en la retina. Cerca de 130 millones de fotorreceptores absorben la luz, sin embargo, aproximadamente 1.2 millones de axones de las células ganglionares transmiten información de la retina al cerebro. El procesamiento en la retina incluye la formación del centro receptivo de células bipolares y ganglionares en la retina, al igual que converge y diverge de las fotorreceptores a la célula bipolar. Además, otras neuronas en la retina, en particular horizontal y células amacrinas, transmiten información lateralmente ( de la neurona en una capa a la neurona adyacente en la misma capa), resultando en un campos receptivos más complejos que pueden ser indiferentes al color y sensibles al movimiento o sensibles al color e indiferentes al movimiento.[24]
Mecanismo de la generación de signos visuales
La retina se adapta para cambiar en la luz a través del uso de los bastones. En la oscuridad, la retina cromofora tiene una forma doblada llamada cis-retinal (refiriéndose a una conformación cis en uno de los enlaces dobles). Cuando la luz interactúa con la retina, cambia la conformación a una forma recta llamada trans-retinal y rompe con la opsina. A esto se le llama blanqueo porque la rodopsina purificada cambia de violeta a incoloro en la luz. En la línea base en la oscuridad, la rodopsina no absorbe luz y libera glutamato el cual inhibe la célula bipolar. Esto inhibe la liberación de neurotransmisores desde las células bipolares hasta la célula ganglionar. Cuando hay luz presente, la secreción de glutamato cesa por lo tanto ya no inhibe a la célula bipolar de liberar neurotransmisores a la célula ganglionar y por lo tanto una imagen puede ser detectada.[25][26]
El resultado final de todo este procesamiento son cinco diferentes poblaciones de células ganglionares que envían información (formación de imágenes y sin formación de imágenes) visual al cerebro:
- Células M, con grandes campos receptivos de envolvente central que son sensibles a la profundidad, indiferentes al color y se adaptan rápidamente a un estímulo.
- Células P, con campos receptivos más pequeños de envolvente central que son sensibles al color y la forma.
- Células K, con grandes campos receptivos de centro solamente que son sensibles al color e indiferentes a la forma o profundidad;
- otra población que es intrínsecamente fotosensible; y
- una población final que se utiliza para los movimientos oculares.[24]
Un estudio de la Universidad de Pensilvania del 2006 calculó el ancho de banda aproximado de las retinas humanas de estar alrededor de 8960 kilobits por segundo, mientras que las retinas de conejillos de india transferían aproximadamente 875 kilobits.[27]
En 2007 Zaidi y sus co-investigadores en ambos lados del Atlántico estudiando a pacientes sin bastones y conos, descubrieron que la nueva célula fotorreceptora ganglionar en humanos también tiene un rol en la visión perceptiva consciente e inconsciente.[28] El pico del espectro sensible fue de 481 nm, Esto demuestra que hay dos caminos para la visión de la retina - una basada en los fotorreceptores clásicos (bastones y conos) y la otra, recién descubierta, basada en célula fotorreceptora ganglionar que actúan como detectores de brillo visual rudimentarios.
Fotoquímica
El funcionamiento de una cámara es a menudo comparado con el funcionamiento del ojo, principalmente porque ambos enfocan la luz de objetos externos en el campo visual sobre un medio sensible a la luz. En el caso de una cámara, este medio es el cine o un sensor electrónico; en el caso del ojo, es una matriz de receptores visuales. Con esta similitud geométrica simple, basada en las leyes de la óptica, el ojo funciona como un transductor, al igual que una cámara CCD.
En el sistema visual, la molécula retinal es una molécula sensora de la luz fundada en las varillas y coníferas de la retina. La molécula retinal es una estructura fundamental que se encarga de la interpretación de la luz hacia las señales visuales, el nervio i.e. empuja hacia el sistema ocular del sistema nervioso central. Cuando se presenta luz, la molécula retinal se configura como resultado de los impulsos nerviosos que son generados.[24]
Nervio óptico
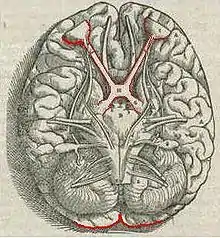
La información acerca de la imagen que es captada por el ojo se transmite al cerebro por medio del nervio óptico. Diferentes poblaciones de células ganglionares en la retina envían información al cerebro a través del nervio óptico. Cerca de un 90% del axón en el nervio óptico pasa por el núcleo glangunado lateral en el tálamo. Estos axones se originan de las células ganglionares de la retina M, P y K. Este proceso paralelo es importante para reconstruir el mundo visual; cada tipo de información pasa por diferentes rutas a para que se dé la percepción. Otra población envía información al colículo superior en el cerebro medio, el cual asiste en controlar el movimiento ocular (movimientos sádicos)[29] al igual que otras respuestas motoras.
Al final la población de células fotosensibles ganglionares, que contienen melanopsina por fotosensibilidad, envían información por medio del tracto retinohipotalámico al pretectum (reflejos pupilares), a varias estructuras involucradas en el control del ritmo circadiano y sueño tal como el núcleo supraquiasmático (el reloj biológico), y hacia el núcleo ventrolateral preóptico (una región involucrada en la regulación del sueño).[30] Un rol recientemente descubierto para las células fotorreceptoras ganglionares es que ellas median la visión consciente e inconscientes - actuando como detectores de brillo visual rudamententario como se muestra en ojos sin bastones ni conos.[28]
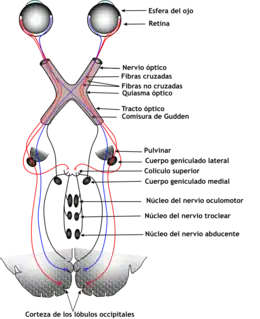
Quiasma óptico
Los nervios ópticos de ambos ojos se encuentran y cruzan en el quiasma óptico,[31][32] en la base del hipotálamo del cerebro. En este punto la información que vienen de los dos ojos es combinado y luego se divide dependiendo del campo visual. Las mitades correspondientes de campo visual (derecho y izquierdo) son enviados a la mitad derecha e izquierda del cerebro respectivamente, a ser procesada. Eso quiere decir, el lado derecho de la corteza visual primaria se encarga con la mitad izquierda del campo visual de los dos ojos, y similar con la mitad izquierda.[29] Una pequeña región en el centro del campo visual es procesado excesivamente por las dos mitades del cerebro.
Tracto óptico
La información del campo visual derecho (ahora en el lado izquierdo del cerebro) viaja en el tracto óptico izquierdo. La información del campo visual izquierdo viaja en el tracto óptico derecho. Cada tracto óptico termina en el núcleo lateral geniculado (NLG) en el tálamo.
Núcleo geniculado lateral
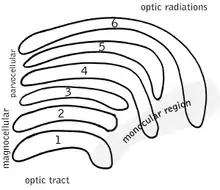
El núcleo geniculado lateral (NGL) se localiza en el tálamo del cerebro. El NGL consiste de seis capas en humanos y otros primates empezando por el catarrhini, incluyendo cercopithecidae y monos. Capas 1, 4 y 6 corresponden a la información de las fibras contralateral (cruzada) de la retina nasal (campo visual temporal); capas 2, 3 y 5 corresponden a la información de las fibras ipsilaterales (no cruzadas) de la retina temporal (campo visual nasal). La capa 1 contiene células M que corresponden a las células M (magnocelular) del nervio óptico del ojo opuesto y se encargan de la percepción de profundidad y el movimiento. Capas 4 y 6 del NGL también se conectan con el ojo opuesto, pero a la célula P (color y bordes) del nervio óptico.En contraste, capas 2, 3 y 5 del NGL conectan a las células M y P (parvocelular) del nervio óptico por la misma parte del cerebro como es respectivo el NGL. Extendido, las seis capas del NGL son el área de una tarjeta de crédito y como tres veces su grosor. El NGL está enrollado en dos elipsoides del tamaño de dos huevos de pájaros. En medio de las seis capas hay pequeñas células que reciben información de la célula K (color) en la retina. Las neuronas del NGL después relevan la imagen visual del córtex visual primario (V1) que está localizado en la parte de atrás del cerebro en el lóbulo occipital y cerca del surco de calcarina. El NGL es no solo una estación de relevo, si no también un centro de proceso; recibe información recíproca de las capas corticales y subcorticales e inervación recíproca del córtex visual.[24]

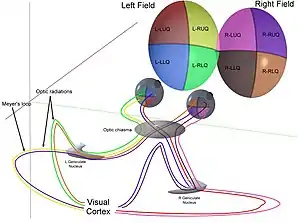
Radiación óptica
La radiación óptica, cada uno de los lados del cerebro cargan información del núcleo geniculado lateral del tálamo a la capa 4 del córtex visual. La capa P de neuronas del NGL releva a la capa V1 4C β. La capa M de neuronas relevan a la capa V1 4C α. La capa K de neuronas del NGL releva a grandes neuronas llamados blobs en la capa 2 y 3 de V1.[24]
Hay una correspondencia directa de una posición angular en el campo visual del ojo, todo el camino por el tracto óptico a una posición de un nervio en V1 ( hasta el V4, i.e. de las áreas visuales primarias. Después de esto, el camino visual es separado hacia el camino dorsal y ventral).
Córtex visual
El córtex visual es el sistema más largo en el cerebro humano y es responsable de procesar una imagen visual. Se encuentra en la parte trasera del cerebro (resaltado en la imagen), por encima del cerebelo. La región que recibe información directamente del NGL se llama el córtex visual primario, (también llamado V1 y córtex estriado). La información visual luego fluye a través de una jerarquía cortical. Estas áreas incluyen V2, V3, V4 y el área V5/MT (la conectividad exacta depende de la especie del animal). Estas áreas visuales secundarias (denominada colectivamente la corteza visual extraestriada) procesan una amplia variedad de primitivas visuales. Las neuronas en V1 y V2 responden selectivamente a barras de orientaciones específicas o combinaciones de barras. Se cree que estas apoyan a la detección de bordes y esquinas. Del mismo modo, la información básica sobre el color y movimiento se procesa aquí.[33]
Heider, et al. (2002) han encontrado que las neuronas que involucran V1, V2 y V3 pueden detectar contornos ilusorios estereoscópicos; estos encuentran que los estímulos estereoscópicos subtendiendo hasta 8º pueden activar estas neuronas.[34]
Córtex de asociación visual
Mientras la información visual pasa a través de la jerarquía visual, la complejidad de las representaciones neuronales aumentan. Las neuronas V1 responden selectivamente a un segmento lineal de una orientación particular en una ubicación retinotópica, las neuronas del complejo lateral occipital responden selectivamente a un objeto completo (e.g., una figura dibujada), y las neuronas en una asociación visual con el córtex responden selectivamente a las caras humanas, o a un objeto particular.
Junto con esta creciente complejidad de representación neural puede venir un nivel de especialización del proceso hacia dos caminos distintos: la corriente dorsal y la corriente ventral[35] (La hipótesis de las dos corrientes, primero propuesta por Ungerleider y Mishkin en 1982). La corriente dorsal, comúnmente referida como la corriente “donde”, está involucrada con la atención espacial (encubierta y abierta), y se comunica con las regiones que controla el movimiento ocular y el movimiento de las manos. Más recientemente, esta área ha sido llamada la corriente “cómo” para enfatizar su rol en conductas de guía en ubicaciones espaciales. La corriente ventral, comúnmente referida como la corriente “qué”, está involucrada en el reconocimiento, identificación y categorización del estímulo visual.

Sin embargo, aún hay mucho que debatir acerca del grado de especialización dentro de estas dos vías, ya que, de hecho, están muy interconectadas.[36]
Horace Barlow propuso la hipótesis de codificación eficiente en 1961 como un modelo teorético de la codificación sensorial en el cerebro.[37]
La red de modo predeterminado es una red de regiones cerebrales que están activas cuando alguien está despierto o dormido. El sistema visual del modo predeterminado puede ser monitoreado durante el estado inactivo del fMRI: Fox, et al. (2005) encontró que “El cerebro humano está organizado por una dinámica de redes anti-correlacionadas funcionales.[38]
En el lóbulo parietal, el córtex intraparietal lateral y ventral están involucrados en la atención visual y los movimientos sacádicos oculares. Estas regiones se encuentran en el surco intraparietal (marcadas en rojo en la imagen adyacente).
Referencias
- "How the Human Eye Sees." WebMD. Ed. Alan Kozarsky. WebMD, 3 October 2015. Web. 27 March 2016.
- Than, Ker. "How the Human Eye Works." LiveScience. TechMedia Network, 10 February 2010. Web. 27 March 2016.
- "How the Human Eye Works | Cornea Layers/Role | Light Rays." NKCF. The Gavin Herbert Eye Institute. Web. 27 March 2016.
- Albertine, Kurt. Barron’s Anatomy Flash Cards
- Tillotson, Joanne. McCann, Stephanie. Kaplan’s Medical Flashcards. April 2, 2013.
- "Optic Chiasma." Optic Chiasm Function, Anatomy & Definition. Healthline Medical Team, 9 March 2015. Web. 27 March 2016.
- Jefferey, G., and M. M. Neveu. "Chiasm Formation in Man Is Fundamentally Different from That in the Mouse." Nature.com. Nature Publishing Group, 21 March 2007. Web. 27 March 2016.
- Card, J. Patrick, and Robert Y. Moore. "Organization of Lateral Geniculate-hypothalamic Connections in the Rat." Wiley Online Library. 1 June. 1989. Web. 27 March 2016.
- Murphy, Penelope C., Simon G. Duckett, and Adam M. Sillito. "Feedback Connections to the Lateral Geniculate Nucleus and Cortical Response Properties." Feedback Connections to the Lateral Geniculate Nucleus and Cortical Response Properties. 19 November 1999. Web. 27 March 2016.
- Schiller, P. H., and J. G. Malpeli. "Functional Specificity of Lateral Geniculate Nucleus Laminae of the Rhesus Monkey." APS Journals. 1 May 1978. Web. 27 March 2016.
- Singer, W., and F. Schmielau. "The Role of Visual Cortex for Binocular Interactions in the Cat Lateral Geniculate Nucleus." The Role of Visual Cortex for Binocular Interactions in the Cat Lateral Geniculate Nucleus. 21 January 1977. Web. 27 March 2016.
- Reed, R. Clay, and Jose-Manuel Alonso. "Specificity of Monosynaptic Connections from Thalamus to Visual Cortex." Letters to Nature. Nature Publishing Group, 3 October 1995. Web. 27 March 2016.
- Heim, Stefan, Simon B. Eickhoff, et al. "Effective Connectivity of the Left BA 44, BA 45, and Inferior Temporal Gyrus during Lexical and Phonological Decisions Identified with DCM." Wiley Online Library. 19 December 2007. Web. 27 March 2016.
- Catani, Marco, and Derek K. Jones. "Brain." Occipito‐temporal Connections in the Human Brain. 23 June 2003. Web. 27 March 2016.
- Benevento, Louis A., and Gregg P. Strandage. "The Organization of Projections of the Retinorecipient and Nonretinorecipient Nuclei of the Pretectal Complex and Layers of the Superior Colliculus to the Lateral Pulvinar and Medial Pulvinar in the Macaque Monkey." Science Direct. 1 July 1983. Web. 27 March 2016.
- Hirsch, JA, and CD Gilbert. "The Journal of NeuroscienceSociety for Neuroscience." Synaptic Physiology of Horizontal Connections in the Cat's Visual Cortex. 1 June 1991. Web. 27 March 2016.
- Schall, JD, A. Morel, DJ King, and J. Bullier. "The Journal of NeuroscienceSociety for Neuroscience." Topography of Visual Cortex Connections with Frontal Eye Field in Macaque: Convergence and Segregation of Processing Streams. 1 June 1995. Web. 27 March 2016.
- Moser, May-Britt, and Edvard I. Moser. "Functional Differentiation in the Hippocampus." Wiley Online Library. 1998. Web. 27 March 2016.
- Kanaseki, T., and J. M. Sprague. "Anatomical Organization of Pretectal Nuclei and Tectal Laminae in the Cat." Anatomical Organization of Pretectal Nuclei and Tectal Laminae in the Cat. 1 December 1974. Web. 27 March 2016.
- Reiner, Anton, and Harvey J. Karten. "Parasympathetic Ocular Control — Functional Subdivisions and Circuitry of the Avian Nucleus of Edinger-Westphal."Science Direct. 1983. Web. 27 March 2016.
- Welsh, David K., and Diomedes E. Logothetis. "Individual Neurons Dissociated from Rat Suprachiasmatic Nucleus Express Independently Phased Circadian Firing Rhythms." Science Direct. Harvard University, April 1995. Web. 27 March 2016.
- Güler, A.D. (May 2008). «Melanopsin cells are the principal conduits for rod/cone input to non-image forming vision» (Abstract). Nature 453 (7191): 102-5. Bibcode:2008Natur.453..102G. PMC 2871301. PMID 18432195. doi:10.1038/nature06829.
- Tovée, 2008
- Saladin, Kenneth D. Anatomy & Physiology: The Unity of Form and Function. 5th ed. New York: McGraw-Hill, 2010.
- «Copia archivada». Archivado desde el original el 23 de enero de 2011. Consultado el 23 de octubre de 2018.
- «Calculating the speed of sight».
- Zaidi FH, Hull JT, Peirson SN (diciembre de 2007). «Short-wavelength light sensitivity of circadian, pupillary, and visual awareness in humans lacking an outer retina». Curr. Biol. 17 (24): 2122-8. PMC 2151130. PMID 18082405. doi:10.1016/j.cub.2007.11.034.
- Sundsten, John W.; Nolte, John (2001). The human brain: an introduction to its functional anatomy. St. Louis: Mosby. pp. 410-447. ISBN 0-323-01320-1. OCLC 47892833.
- Lucas RJ, Hattar S, Takao M, Berson DM, Foster RG, Yau KW (enero de 2003). «Diminished pupillary light reflex at high irradiances in melanopsin-knockout mice». Science 299 (5604): 245-7. Bibcode:2003Sci...299..245L. PMID 12522249. doi:10.1126/science.1077293.
- Turner, Howard R. (1997). «Optics». Science in medieval Islam: an illustrated introduction. Austin: University of Texas Press. p. 197. ISBN 0-292-78149-0. OCLC 440896281.
- Vesalius, 1543
- Jessell, Thomas M.; Kandel, Eric R.; Schwartz, James H. (2000). «27. Central visual pathways». Principles of neural science. New York: McGraw-Hill. pp. 533–540. ISBN 0-8385-7701-6. OCLC 42073108.
- Heider, Barbara; Spillmann, Lothar; Peterhans, Esther (2002). «"Stereoscopic Illusory Contours— Cortical Neuron Responses and Human Perception" ». J. Cognitive Neuroscience 14 (7): 1018-1029. Archivado desde el original el 11 de octubre de 2016. Consultado el 18 de mayo de 2014.
- Mishkin M, Ungerleider LG (1982). «Contribution of striate inputs to the visuospatial functions of parieto-preoccipital cortex in monkeys». Behav. Brain Res. 6 (1): 57-77. PMID 7126325. doi:10.1016/0166-4328(82)90081-X.
- Farivar R. (2009). «Dorsal-ventral integration in object recognition». Brain Res. Rev. 61 (2): 144-53. PMID 19481571. doi:10.1016/j.brainresrev.2009.05.006.
- Barlow, H. (1961) "Possible principles underlying the transformation of sensory messages" in Sensory Communication, MIT Press
- Michael D. Fox, et al., PNAS vol. 102 no. 27 9673–9678, doi:10.1073/pnas.0504136102
 Datos: Q558363
Datos: Q558363 Multimedia: Visual system / Q558363
Multimedia: Visual system / Q558363