Meristemo
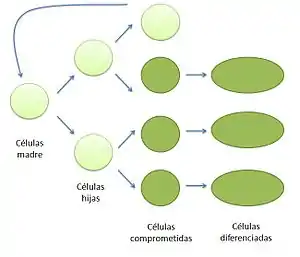
Los meristemos —del griego μεριστός, "divisible"—[1], o tejidos meristemáticos son los responsables del crecimiento vegetal. Sus células son pequeñas, tienen forma poliédrica, paredes finas y vacuolas pequeñas y abundantes. Se caracteriza por mantenerse poco diferenciado. Tienen capacidad de división y de estas células aparecen los demás tejidos. Las principales características de las células meristemáticas son su aspecto poliédrico y equidimensional, la presencia de vacuolas pequeñas y abundantes, así como de una pared celular delgada, sin pared secundaria. Asimismo, se mantienen siempre jóvenes y poco diferenciadas. Estas células son totipotentes, con capacidad para dar lugar a todos los demás tejidos, siendo homólogas a las células madre de los animales. Las células meristemáticas se dividen por mitosis dando lugar a dos células hijas: una que retiene su carácter meristemático indefinidamente y otra que se desarrolla y se especializa en un tejido concreto.
Las plantas, a diferencia de los animales, tienen un sistema abierto de crecimiento. Esto significa que la planta posee regiones embrionarias más o menos perennes, de las cuales se producen periódicamente nuevos tejidos y órganos.

Histología
Los meristemos están compuestas por células no diferenciadas que se dividen activamente, también llamadas células totipotentes por su habilidad de dar lugar a todos los tejidos vegetales. Típicamente, las células meristemáticas son chiquitas poliédricas, más o menos equidimensionales (dimensiones parecidas en todas las direcciones). En ellas, el citoplasma ocupa la mayor parte de volumen celular ya que las vacuolas son muy pequeñas, las células meristemáticas no contienen cloroplastos ni ningún otro plástido diferenciado, la pared celular de las células meristemáticas es delgada y carece de pared secundaria.
En las plantas, las células meristemáticas son homólogas funcionales de las células madre que dan lugar a todos los tejidos en animales. Estas células se dividen en otras dos, con el fin de dar origen a una célula hija que continúa creciendo de forma meristemática, la cual retiene su carácter embrionario indefinidamente y en otra que se diferencia en una vía de desarrollo particular formando muchos tipos de tejidos.[2]
Los meristemos en las plantas pueden ser clasificados de varias formas:
De acuerdo a su origen pueden dividirse en meristemos primarios y secundarios. Los meristemos primarios suelen generar los órganos y tejidos que constituyen el cuerpo de la planta, ejemplos de tales meristemos son los apicales, del tallo y la raíz. Los meristemos secundarios se forman en la mayoría de las plantas y pueden tener estructuras diferenciadas de los meristemos primarios, son ejemplos de estos los meristemos axilares, de inflorescencia, florales, intercalares y laterales.[3][4]
También se pueden clasificar los meristemos de acuerdo a su posición espacial y a los tejidos u órganos que originen. De acuerdo a su posición espacial se los suele referir como meristemos apicales, laterales o intercalares.[2]
Meristemos primarios o apical
Los meristemos primarios se originan en el embrión y son los responsables del crecimiento en longitud de la planta. Los primeros meristemos en aparecer durante el desarrollo del cuerpo vegetativo de una planta vascular están localizados en la punta de tallos y raíces. Debido a su localización, estos meristemos son llamados meristemos apicales. Todos los tejidos meristemáticos primarios y por lo tanto todos los tejidos primarios de la planta se originan a partir del meristemo apical de la raíz o del meristemo apical del brote.
El meristemo apical de la raíz habitualmente está cubierto por una estructura de células diferenciadas que lo protege, conocida como cofia. El meristemo apical del tallo (o yema terminal) puede estar desnudo o cubierto por hojas. En este caso, las hojas son llamadas primordios foliares, que tienen un rudimento de yema auxiliar en su base. Este se convertirá en una yema cuando las hojas se desarrollen, y dará lugar a una nueva rama.
Las células que mantienen al meristemo con un flujo constante de células nuevas son llamadas células iniciales. estas se dividen de tal manera que entre pares de células hermanas una de ellas se convierte en desecho y la otra es capaz de dar origen a un nuevo cuerpo celular, estas últimas se conocen como células derivadas las cuales logran dividirse muchas veces. r a la superficie del meristemo y por ello sus células hijas permanecerán siempre en la misma capa. Las divisiones celulares de la capa L3 tienden a ser menos regulares y en todos los planos, así mismo, son capaces drior del meristemo. Para poder determinar las estructuras que originan estas diferentes capas celulares, investigadores han optado por la construcción de quimeras. Las plantas quiméricas están compuestas de capas que dan origen a fenotipos con marcadores distinguibles, de acuerdo a diferencias en el genotipo de cada capa. Las quimeras han sido usadas además para demostrar la capacidad de las capas de que cada capa celular individual puede influir sobre el desarrollo de una capa adyacente.[2]
Además de la organización por capas, los meristemos apicales activos también poseen un patrón de zonas funcionales de organización radial llamado zonación citohistológica. Cada zona está compuesta por células que pueden ser distinguidas de acuerdo con su plano de división, su tamaño y sus grados de vacualización. Estas zonas también exhiben diferentes patrones de expresión genética, reflejando una diferencia funcional en cada zona. La primera de estas es la zona central, la cual es encontrada en la parte superior de los centros de los meristemos activos y está compuesto por células altamente vacuoladas que se dividen infrecuentemente, es un lugar de autorenovación de células madre. Flanqueando la zona central se encuentra la zona periférica, una región en forma de dona de células más pequeña y con una mayor tasa de división celular las cuales darán lugar a la formación de las hojas y los meristemos florales de inflorescencia. Finalmente se encuentra la zona de costilla, debajo de las dos regiones anteriormente descrita. En esta zona, la diferenciación y elongación celular darán lugar al tejido interno del tallo de la planta.[3]
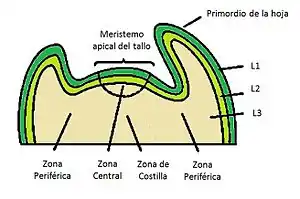
Dentro de los diferentes tejidos de tallos y raíces, los meristemos apicales darán origen a los siguientes tejidos:
- Protodermo: se localiza alrededor y al exterior, da origen a la epidermis.
- Procambium: se localiza al interior del protodermo, da lugar a los tejidos vasculares: xilema, floema y cámbium vascular.
- Meristema fundamental: se localiza entre el Protodermo y Procambium, da origen a parénquima, colénquima y esclerénquima.
- Meristemos remanentes: Actúan cíclicamente. Se localizan en la base de los entrenudos que están quiescentes (latentes).
- Meristemos meristemoides: son células adultas diferenciadas que por ser células vivas tienen la propiedad de poder desdiferenciarse y volver a ser meristemáticas y dividirse por mitosis, originando nuevas estructuras, como células epidérmicas que originan estomas, pelos o tricomas y aguijones, etcétera.
Meristemos laterales o secundarios
Son los responsables del crecimiento radial (secundario). Dan lugar a xilema, floema y parénquima secundario (cámbium) y a parénquima cortical y suber (felógeno), y contribuyen al engrosamiento de tallos y raíces por formación de capas concéntricas nuevas que dan lugar además a un engrosamiento de los ejes. Están distribuidos por toda la planta. Sus células recuperan su capacidad meristemática y comienzan a dividirse formando nuevas células, dando lugar a un crecimiento en grosor en tallos y raíces de plantas leñosas. Los meristemos secundarios son de dos tipos:[3]
- Cámbium vascular: Es un meristemo secundario que se diferencia junto con el tejido vascular primario dentro del cilindro vascular. No produce los órganos laterales, pero sí el tejido leñoso de tallos y raíces. El cámbium vascular posee dos tipos de células meristemáticas dependiendo de la especie de planta: las células madre fusiformes y las células madre de los rayos. Las células madre fusiformes son células alargadas y vacuoladas las cuales se dividen longitudinalmente para regenerarse a sí mismas y cuyos derivados se diferenciarán en las células conductoras del xilema y del floema. Las células madre de los rayos son células pequeñas cuyos derivados incluyen las células parinquematosas orientadas radialmente dentro de la madera.
- Cámbium felógeno: Es una capa de células meristemáticas que se desarrollan entre las células de la corteza y del floema secundario. De estas células se derivan las células de la peridermis, las cuales constituyen una superficie externa protectora del cuerpo secundario de la planta, reemplazando la epidermis por una corteza en las ramas.
Meristemos intercalares
Los tallos de las monocotiledóneas sí tienen meristemos secundarios laterales; sin embargo, frecuentemente tienen meristemos intercalares insertados en los tallos entre los tejidos maduros aumentando su longitud. Se encuentran por lo general en la base de los tallos o de las hierbas, debido a este tipo de meristemos, el césped puede seguir creciendo después de ser cortado.[3]
Diferenciación del meristemo
Una característica de las células meristemáticas es su posición con respecto a otras células y esto tiene que ver con su geometría y aunque parece tener un papel importante en la determinación de las naturaleza de la división de los meristemos, las sustancias químicas tienen una influencia más fuerte. La razón Citoquinina/Auxina regula la producción de raíces y tallos por parte de las células del meristemo. Aparentemente las células indiferenciadas de las plantas tienen dos opciones: pueden elongarse o dividirse repetitivamente. Las células que se dividen repetitivamente permanecen indiferenciadas o meristemáticas mientras que las que se elongan son principalmente diferenciadas. Autores experimentaron con la planta de Tabaco (Nicotiana sp.) adhiriendo Ácido Indoleacético(IAA) a los tejidos del tallo esto produjo la expansión rápida de las células formando de este modo células gigantes. La Kinetina sola por ejemplo, tiene muy poco efecto sobre las células, pero el IAA más la Kinetina hace que las células se dividan muy rápido. El resultado es un gran número de células relativamente pequeñas no diferenciadas. En otras palabras las células permanecen meristemáticas en ciertas concentraciones de Auxina/Citoquinina. En otros caso se ha probado que altas concentraciones de Auxina en el 'tejido calloso' dan lugar a la formación de raíces, por su parte la kinetina más la auxina pueden determinar no solo el lugar donde se formaran las raíces si no también las yemas apicales.
Véase también
Referencias
- http://lema.rae.es/drae/?val=meristemo
- Gilbert, Scott F (2006). Biología del Desarrollo. Buenos Aires, Argentina: Editorial Médica Panamericana.
- Taiz, Lincoln (2007). Plant Physiology. Sinauer Associates Inc. Publishers. ISBN 978-0-87893-866-7.
- P. Font Quer - Diccionario de Botánica - 1982
https://www.innovabiologia.com/biodiversidad/diversidad-vegetal/los-meristemos/
Bibliografía
- Neil A. Campbell and Jane B. Reece Biology, 6th edition, Benjamin Cummings.
- Frank S. & Robert V. Las Plantas Vasculares: Forma y Función
- Carlos Andres Casado Crispin estudio en el colegio INESAM científico de Biology of Plants, 7th edition, 2004.