Poliploidía
En genética, la poliploidía se define como el fenómeno por el cual se originan células, tejidos u organismos con tres o más juegos completos de cromosomas de la misma o distintas especies o con dos o más genomas de especies distintas. Dichas células, tejidos u organismos se denominan poliploides. Es uno de los pocos casos de saltacionismo confirmados.
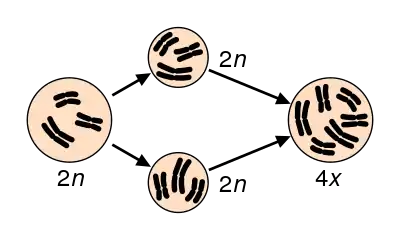
Si los genomas de una especie poliploide provienen de la misma especie ancestral, se dice que es autopoliploide o autoploide; si provienen de dos especies ancestrales diferentes, se dice que es alopoliploide o aloploide. Dependiendo del número de juegos cromosómicos completos que posee la especie se denomina triploide (3n), tetraploide (4n), pentaploide (5n), hexaploide (6n) y así sucesivamente, donde n es el número haploide, y no hay que confundirlo con el número monoploide (x).
Los elementos genéticos redundantes de los organismos poliploides pueden llegar a transformarse en copias únicas mediante mecanismos de subfuncionalización, neofuncionalización o mediante la aparición de determinadas mutaciones.
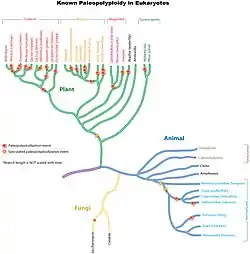
La poliploidía es un suceso bastante frecuente en la naturaleza, aunque es más común en plantas[1] y algas que en animales y hongos. En plantas, la poliploidía se encuentra muy extendida dentro de las angiospermas (aproximadamente un 30% de las especies son alopoliploides) y parece estar relacionada con la latitud geográfica. Generalmente en las plantas poliploides se da el fenotipo giga: se produce un aumento de tamaño en los individuos poliploides, ya que poseen mayor número de células que los individuos los diploides. En animales, el fenotipo giga no se da como tal, ya que el aumento de tamaño que se da en algunos grupos de insectos, crustáceos y algunos anfibios y peces poliploides se debe al aumento del volumen celular pero no del número de células, siendo este último el mismo en individuos diploides y poliploides.
Tipos
De acuerdo con el número de juegos de cromosomas que existen en el núcleo de la célula, pueden establecerse los siguientes grupos de poliploidía:
- Triploide (3 juegos de cromosomas).
- Tetraploide (4 juegos de cromosomas).
- Pentaploide (5 juegos de cromosomas).
- Hexaploide (6 juegos de cromosomas).
- Heptaploide o septaploide (7 juegos de cromosomas).
- Octaploide (8 juegos de cromosomas).
- Nonaploide (9 juegos de cromosomas).
- Decaploide (10 juegos de cromosomas).
- Dodecaploide (12 juegos de cromosomas).
Terminología
A continuación se definen algunos términos frecuentemente utilizados en el estudio de poliploides:
Diploide (2X): especie o individuo cuya dotación autosómica normal está compuesta por dos juegos idénticos de X cromosomas cada uno, siendo cada uno de estos X cromosomas diferentes entre sí y no agrupables en grupos inferiores que a su vez constituyeran juegos cromosómicos de especies parentales.
Número monoploide o número básico (X): número de cromosomas diferentes que constituyen un juego cromosómico completo.
Número haploide (n): número de cromosomas que se incluyen en un gameto normal de una especie.
Poliploide: especie o individuo cuya dotación cromosómica es un múltiplo de X mayor que 2X.
Autopoliploide o autoploide: poliploide que presenta un solo tipo de genoma y, por tanto, procede de una única especie parental.
Alopoliploide o aloploide: poliploide que presenta más de un tipo de genoma y, por tanto, procede de la hibridación de dos o más especies. En ocasiones, se desconoce el origen de alguno de los genomas de una especie aloploide, pudiendo pertenecer a una especie parental extinta o no descubierta.
Alopoliploide segmentado o aloploide segmentado: poliploide que presenta algún juego cromosómico incompleto, pero que suple la carencia cromosómica de dicho juego con la aneuploidía de los homeólogos que sí están presentes, siendo el número de copias extras igual al número de homeólogos ausentes. Esto puede ocurrir con segmentos cromosómicos en lugar de cromosomas completos o ambos cosas simultáneamente. También son alopoliploides segmentados aquellos poliploides que presentan diferente número de juegos cromosómicos para cada genoma parental.
Mixoploide: individuo que presenta líneas celulares con distinto nivel de ploidía.
Poliploidía en la naturaleza
La poliploidía, en la naturaleza, es un fenómeno espontáneo que se da tanto en animales como en plantas, si bien es mucho más frecuente en estas últimas.
En plantas
La poliploidía se encuentra en todos los grandes grupos vegetales.
La distribución geográfica de las especies vegetales muestra un incremento de la proporción de especies poliploides a medida que aumenta la latitud, siendo la explicación más plausible la mayor adaptabilidad de los poliploides a condiciones climáticas como el fotoperiodo, si bien existe cierta controversia en la correlación entre poliploidía y resistencia a condiciones adversas. Según Gustafsson (1948), la elevada proporción de poliploides en la flora ártica no es debida a la acción de la temperatura extrema sobre la gametogénesis, sino a adaptaciones en los ciclos vitales de estos organismos. Cabe destacar que existe una tendencia a una distribución más amplia entre los poliploides y una diferenciación en los hábitats entre los poliploides y los diploides a partir de los que se originaron.
Existe bastante controversia sobre si la autopoliploidía ha jugado un papel importante en la evolución de las especies vegetales o no, pero no cabe duda de que la alopoliploidía ha tenido un papel destacable en la evolución de estos grupos. Winge (1917) fue el primero en resaltar la importancia de la hibridación interespecífica y posterior duplicación cromosómica en la formación de nuevas especies. Esta hipótesis está apoyada por el experimento llevado a cabo por Clausen y Goodspeed (1925), en el que se obtuvo una nueva especie, Nicotiana digluta (2n = 72), a partir de N. glutinosa (2n = 24) y N. tabacum (2n = 48).
La alopoliploidía constituye un mecanismo evolutivo discontinuo, ya que reúne los patrimonios hereditarios de especies distintas al producirse la hibridación y la duplicación cromosómica en una sola generación. Aunque las etapas iniciales son cruciales para el establecimiento y la supervivencia de los poliploides, una vez superadas pueden extenderse por nuevas áreas geográficas. Además, sus irregularidades meióticas pueden ser compensadas por mecanismos apomícticos. Después de la formación de una especie poliploide, esta evoluciona lentamente debido a que la variación fenotípica es menor que en diploides, ya que un mutante recesivo tiene menos probabilidad de manifestarse y, por tanto, de que pueda actuar, tanto beneficiosa como negativamente, la selección sobre él. Es común que entre los subgenomas de la especie poliploide se produzcan reordenamientos intercromosómicos.
A pesar de los numerosos intentos en los últimos 70 años para estimar frecuencias de poliploidía en plantas, ha sido dificultoso determinar la frecuencia real en numerosos linajes de plantas. Las angiospermas en particular han recibido mucha atención. Utilizando los números cromosómicos e hipótesis acerca de la presunta línea divisoria entre el número diploide y poliploide, se han realizado varias estimaciones de la frecuencia de la poliploidía en las plantas con flores. Sin embargo, tal estimación ha variado dependiendo del número cromosómico básico utilizado, como así también de los taxones utilizados como muestra. Arne Müntzing, en 1936,[2] y Cyril Dean Darlington, en 1937,[3] especularon que cerca de la mitad de las angiospermas eran poliploides, mientras que George Ledyard Stebbins, en 1950,[4] estimó tal frecuencia en 30 a 35 %. Verne Grant basó su estimación en el número cromosómico para 17 138 especies e hipotetizó que todas las que tenían un número haploide superior a n=14 eran poliploides. Basado en estos parámetros concluyó que el 47 % de las angiospermas (58 % de las monocotiledóneas y 43 % de las dicotiledóneas) eran poliploides.[5] Peter Goldblatt sugirió que tal estimación era muy conservadora y postuló que todos los taxones con números cromosómicos básicos por encima de x=8 o x=9 debían tener un origen poliploide. Utilizando esta regla, calculó que el 70% de las monocotiledóneas eran poliploides. Lewis utilizó similar razonamiento para las dicotiledóneas, llegando a estimar que el 70 a 80 % de las mismas eran poliploides.[6]
En animales
Según White (1951), los insectos y crustáceos partenogenéticos presentan razas y especies poliploides. Se han estudiado multitud de series poliploides, encontrándose en todas ellas una estrecha relación con la partenogénesis: Artemia salina, Trichiniscus elizabethae, Solenobio triguetrella, Oetiorrhycus sp., etc.
Muller (1925) atribuyó la baja frecuencia de poliploidía en animales superiores al desequilibrio que origina el incremento del número de cromosomas sexuales en el mecanismo genético de determinación del sexo. Basó esta argumentación en la determinación sexual de Drosophila melanogaster en diferentes estados de ploidía, basándose en los conocimientos de la época:
- X/A = 1: hembra.
- X/A = 0.5: macho.
- 1 > X/A > 0.5: intersexo (estéril).
Siendo A un autosoma y X el cromosoma X. En un individuo triploide:
- AAA XXX: X/A = 1: hembra.
- AAA XXY: X/A = 0.66: intersexo (infértil).
- AAA XYY: X/A = 0.33: supermacho (infértil).
En un individuo tetraploide:
- AAAA XXXX: X/A = 1: hembra.
- AAAA XXXY: X/A = 0.75: intersexo (infértil).
- AAAA XXYY: X/A = 0.5: macho.
Debido a la formación de bivalentes XX e YY en la meiosis de los machos tetraploides, dos tercios de sus gametos serán XY y, por consiguiente, la mayor parte de la descendencia será XXXY (intersexo).
Dawson (1962) sugirió otras dos razones importantes para la baja frecuencia de poliploidía entre los animales:
- El origen de una raza tetraploide a partir de una diploide requiere la ocurrencia de dos poliploidizaciones: una de un macho y una de una hembra, así como su posterior apareamiento, siendo muy improbable que esto ocurra.
- El desequilibrio genético por la poliploidía en los procesos de diferenciación de los tejidos y su interacción, lo que causa infertilidad o inviabilidad en muchos casos.
Dentro de los animales poliploides, podemos distinguir aquellos cuya reproducción es partenogenética o asexual de los que presentan una reproducción bisexual normal. Antes se han citado algunos ejemplos de animales poliploides con reproducción partenogenética. Entre las especies animales poliploides bisexuales merece la pena destacar a las ranas del Brasil de la familia Ceratophrididae, en las que Odontophrinus cultripes es diploide (2n = 22), O. americanus es tetraploide (2n = 44) y Ceratophrys dorsata es octoploide (2n = 104).
Según Astaurov (1969) para que se establezca una especie poliploide bisexual primero debe producirse una línea poliploide partenogenética y después debe ocurrir una reversión a la bisexualidad.
Beçak y colaboradores (1970) proponen que en peces y en anfibios la formación de nuevas especies por poliploidía sigue siendo un mecanismo evolutivo importante debido a que los cromosomas sexuales de estas especies están en un estado indiferenciado.
La poliploidía es un fenómeno poco frecuente en animales. Se conocen especies poliploides en anélidos, crustáceos, anfibios y reptiles. En peces, se ha demostrado la existencia de poliploides naturales en algunas especies.[7] Especies de tetraploides ancestrales con aproximadamente 100 cromosomas se han encontrado en los géneros Carassius, Cyprinus, Spinibarbus y Tor mientras que a Schizothorax taliensis y S. yunnanensis son hexaploides ancestrales con 148 cromosomas. En los casos de Carassius auratus y Carassius gibelio además de ser considerados como tetraploides ancestrales, también presentan formas triploides naturales con aproximadamente 150 cromosomas.[8][9][10]
La poliploidía es considerada un carácter inusual en moluscos bivalvos[11] Sin embargo, la evidencia citogenética indica que las especies de los géneros dulceacuícolas Sphaerium, Musculium y Pisidium, adscritos a la subfamilia Sphaeriinae (Sphaeriidae), serían poliploides. Se han documentado antecedentes citogenéticos para 10 especies de Sphaeriinae que habitan en el hemisferio norte, las cuales se caracterizan por presentar los más altos niveles de ploidía descritos dentro de la clase Bivalvia, con valores desde 2n hasta 13n y con números cromosómicos que varían desde 36 hasta ca. 247.[12][13][14]
Identificación
Para tener la certeza de que un individuo es poliploide es necesario el uso de técnicas citogenéticas de estudio cromosómico, como bandeos cromosómicos, Hibridación fluorescente in situ (FISH) y la observación del apareamiento meiótico. Pese a ello, algunas características morfológicas, citológicas y fisiológicas pueden ser útiles a la hora de discriminar entre individuos diploides y poliploides.
En lo referente a las características morfológicas, estas difieren entre plantas y animales. A menudo, en plantas se produce gigantismo (fenotipo giga), siendo todos los órganos de la planta poliploide de mayor tamaño y además el color de las hojas se torna más oscuro. Sin embargo, se admite la existencia de un grado de ploidía óptimo a partir del cual un incremento en el número de juegos cromosómicos no produce un incremento del tamaño, sino que este no varía o, incluso, disminuye. En animales, no se da el fenotipo giga; en anfibios, los individuos triploides obtenidos son del mismo tamaño que los diploides, pues al aumentar el tamaño celular con la ploidía también se produce una disminución del número de células en diversos órganos. Esta disminución del número de células en cerebro y espina dorsal es la causa de trastornos fisiológicos y metabólicos que suelen producir la inviabilidad de los individuos poliploides. Pese a esto, en algunos invertebrados la poliploidía va asociada a un aumento de tamaño.
En cuanto a características citológicas, en multitud de especies vegetales y animales se ha observado una estrecha correlación entre la ploidía de las células y el tamaño unidimensional y tridimensional de las mismas. Además, el aumento del tamaño celular se expresa como una disminución del número de núcleos por unidad de superficie. Una característica citológica que a menudo se ha utilizado en la identificación de poliploides animales es la relación entre el número máximo de nucleolos y el número de juegos cromosómicos presentes en el núcleo.
En relación con las características fisilógicas, en general se admite que el desarrollo ontogenético en poliploides es más lento que diploides, tanto en plantas como en animales.
Citogenética
Mitosis
La mitosis de los individuos poliploides, en general, no presenta ningún tipo de irregularidad que la distinga de la mitosis de los individuos diploides, razón por lo cual la reproducción asexual está tan extendida entre los poliploides.
Meiosis
Dado su diferente origen, los comportamientos meióticos entre autopoliploides y alopoliploides también muestran diferencias, por lo que se tratarán de manera independiente en los siguientes apartados.
Autopoliploides
Entre los autopoliploides, podemos a su vez distinguir entre autotriploides y autotetraploides. Aunque caben otras posibilidades, se tratarán estás por ser las más comunes y las de mayor interés.
Autotriploides
Presentan tres copias de cada cromosoma.
En la profase meiótica el apareamiento entre regiones homólogas de los cromosomas puede ser múltiple o, por el contrario, dos o dos, siendo más frecuente este último tipo.
Al haber más de dos cromosomas homólogos y siendo más frecuente el apareamiento dos o dos, pueden originarse cambios de pareja en el apareamiento, dando lugar a la formación de multivalentes en la profase I. Estos quedan unidos por quiasmas, adoptando múltiples configuraciones metafásicas en función del lugar donde ocurran los quiasmas:
- Cadena: los quiasmas se dan entre los extremos del multivalente dispuesto de manera lineal.
- Y griega: los quiasmas unen a los tres cromosomas por un único extremo.
- Sartén: dos cromosomas quedan unidos por quiasmas en sus dos extremos y el otro queda unido externamente a uno de los extremos de la pareja anterior.
- Parrilla: el tercer cromosoma queda entre los otros dos y los tres se unen por ambos extremos.
También puede ocurrir que el apareamiento se dé entre dos cromosomas, formando un bivalente, y el tercero quede sin aparear (univalente). Que se dé esta situación o se formen trivalentes depende de los siguientes factores:
- La longitud de los cromosomas: a mayor longitud, mayor probabilidad de que se asocien los tres cromosomas.
- El número total de cromosomas en el núcleo: cuanto mayor es el número, mayor es la interferencia entre cromosomas y más difícil es la formación de trivalentes.
- La frecuencia de quiasmas: cuanta mayor frecuencia, mayor será la probabilidad de formación de trivalentes.
Estos factores dependen tanto de la especie como del genotipo del individuo.
En metafase I, los centrómeros de los trivalentes se coorientan, pudiendo ser esta coorientación de tres tipos:
- Lineal: dos cromosomas van a polos opuesto y el tercero queda retrasado en la placa metafásica.
- Convergente: dos cromosomas van hacia un polo y el otro hacia el opuesto.
- Indiferente: puede evolucionar hasta cualquiera de las dos situaciones anteriores.
Los cromosomas que quedan retrasados en la placa metafásica pueden quedar perdidos en el citoplasma o incluirse casualmente en uno de los núcleos hijos.
Autotetraploides
La conducta meiótica de los autotetraploides es muy similar a la descrita para los autotriploides, solo que al ser cuatro los cromosomas homólogos implicados, la situación en un poco más compleja.
En estos individuos, el apareamiento cromosómico puede dar lugar a un tetravalente, un trivalente y un univalente, dos bivalentes o un bivalente y dos univalentes. Se prestará atención al caso de los tetravalentes. En el caso de los tetravalentes, existe un mayor número de asociaciones posibles, diez según Darlington (1931), pero no todas ellas tienen la misma frecuencia, destacando especialmente las asociaciones en anillo y las lineales.
En metafase I, los coorientación de los centrómeros puede presentar cuatro tipos diferentes:
- Lineal: dos cromosomas segregan hacia polos opuestos y los otros dos quedan retrasados en la placa metafásica (pseudounivalentes).
- Convergente: dos cromosomas segregan hacia un polo y los otros dos hacia el otro.
- Paralela: tiene el mismo resultado que la anterior.
- Indiferente: puede evolucionar hacia cualquiera de las situaciones anteriores.
Alopoliploides
El comportamiento meiótico de los alopoliploides está principalmente influido por la afinidad existente entre los genomas de las especies parentales que lo componen.[15]
Si los genomas parentales son muy diferentes, en las células meióticas del alopoliploide solo se observarán bivalente y quizás algún univalente producido por la interferencia de algún cromosoma extraño o por la acción del citoplasma de la especie materna original.
Por el contrario, si los genomas parentales están próximos filogenéticamente, habrá una mayor probabilidad de que se formen multivalentes. Aunque este pueda parecer el hecho más común, depende de lo avanzado que esté el proceso de diploidización del alopoliploide, es decir, como de similar sea el comportamiento meiótico al de los diploides, encontrando cada vez un mayor número de bivalentes y un menor apareamiento entre cromosomas homeólogos. Esta pérdida de afinidad entre los genomas parentales puede ser atribuida al acúmulo de mutaciones estructurales que produzcan una divergencia evolutiva de ambos genomas o bien a alguna alteración en la regulación genética del apareamiento meiótico.
Otras características destacables de la meiosis de alopoliploides son las siguientes:
- Asociaciones secundarias entre bivalentes: los bivalentes metafásicos de cromosomas con alguna relación genética o evolutiva no se distribuyen al azar, sino que forman grupos.
- Fraccionamiento del complemento cromosómico, partición genómica y eliminación de cromosomas.[16]
Fertilidad
La fertilidad de un poliploide depende de la viabilidad de los gametos y de los cigotos que producen, siendo estos a su vez dependientes de la constitución cromosómica, la cual resulta más equilibrada en los autotetraploides que en los autotriploides dado el comportamiento meiótico de los mismos.
Los individuos triploides son prácticamente ésteriles. salvo excepciones como la espinaca.[17] Sin embargo, los tetraploides suelen ser fértiles, aunque su fertilidad es menor que presentada por diploides. Gracias al predominio de las coorientaciones convergente y paralela en Metafase I, la proporción de gametos viables producidos por los tetraploides es mayor que los producidos por los triploides.
En autopoliploides, una selección continuada de individuos tetraploides con genotipos que tienden a formar un mayor número de tetravalentes con coorientación apropiada o un mayor número de bivalentes conlleva un incremento en la fertilidad y en la regularidad meiótica de los individuos de la estirpe tetraploide.
En alopoliploides, la fertilidad puede verse influida por la interacción entre el citoplasma de la especie parental materna y el genoma de la especie parental paterna. Dado que, a priori, una mayor regularidad en la formación de bivalentes meióticos supone una mayor fertilidad en aloploides, cuanto más estéril sea un híbrido, más fértil debe ser el aloploide que de él derive.
Herencia
El tipo de herencia que presentan los poliploides es la llamada herencia poligénica, en la que las segregaciones genotípicas y fenotípicas son diferentes de las mendelianas, dependiendo estas del grado de ploidía, de la frecuencia de los distintos tipos de coorientaciones en metafase y además, en aloploides, de la semejanza genética entre los genomas parentales. Para un locus con dos alelos (A y a), el número de genotipos posibles dependerá del grado de ploidía de las células, es decir, del número de copias de cada gen. En un n-ploide, cada alelo puede estar presente desde 0 hasta n veces. Utilizando la terminología de Belling y col., para un individuo tetraploide se presentan los siguientes genotipos:
- Nuliplexo: aaaa
- Simplexo: Aaaa
- Duplexo: AAaa
- Triplexo: AAAa
- Cuadruplexo: AAAA
Los fenotipos dependerán del cuantas dosis necesite el alelo A para ser dominante. En el caso de que sea dominante a cualquier dosis, el fenotipo de los individuos simplexo, duplexo, triplexo y cuadruplexo será dominante y únicamente el nuliplexo presentará fenotipo recesivo, siendo esta la razón por la que los poliploides evolucionan más lentamente que los diploides, siendo dicha lentitud mayor cuanto mayor es el grado de ploidía.
En el caso de los alopoliploides, Burnham (1962) describió una serie de características:
- Una menor tasa de mutación aparente, al ser la mayoría de la mutaciones recesivas y quedar enmascaradas por los alelos dominantes.
- Una compensación de las funciones génicas, al igual que ocurre con los aneuploides.
- Un tipo de herencia polisómica debido a la repetido de un mismo locus en diferentes genomas parentales.
- Una menor esterilidad de los heterocigotos estructurales en comporación con lo que ocurre en especies diploides.
Usos en Mejora Genética
En animales
Las inducción de la poliploidía no tiene por qué suponer un aumento del tamaño del animal. Además, esta no es aplicable a todas las especies y su obtención a menudo es esporádica, por lo que no resulta rentable económicamente, salvo en el caso de organismos acuáticos como los peces y los molusco.
En Acuicultura, la obtención de individuos poliploides se realiza en huevos recién fecundados por los siguientes procedimientos:
- Choques térmicos, pudiendo ser fríos o calientes.
- Choques de presión hidrostática.
- Utilización de agentes químicos.
Los choques térmicos calientes suelen ser más efectivos que los fríos, siendo así para la trucha arcoíris (Salmo gairdneri), la trucha, la carpa, los peces planos y algunos moluscos, como la ostra japonesa. En cuanto a la técnica de la presión hidrostática, en trucha arcoíris se ha utilizado en la obtención de triploides por fusión del pronúcleo meiótico masculino (n) con el núcleo femenino ameiótico (2n) y en la obtención de tetraploides a partir de la fusión de los núcleos de las células hijas de la primera división meiótica. En lo referente a la utilización de agentes químicos, un método muy utilizado en la obtención de moluscos (ostras y vieiras) triploides y tetraploides consiste en la utilización de citocalasina B.
Aunque en plantas, la poliploidía induce la formación de fenotipos gigas, esto no ocurre en animales, por lo que organismos poliploides no tienen por qué ser de mayor tamaño que los diploides de los que proceden. Aun así, la poliploidización puede ser interesante en la Mejora Genética de animales, ya que los triploides estériles destinan más recursos al crecimiento y, por consiguiente, se obtienen individuos de mayor peso.
Uso de autopoliploides
La fácil inducción de la poliploidía mediante el uso de colchicina, así como el fenómeno de fenotipos gigas que se observa en plantas poliploides podría llevarnos a pensar que la autopoliploidía resulta económicamente beneficioso. Sin embargo, no todas las especies en las que se ha estudiado la autopoliploidía han resultado serlo.
En 1945, Levan propuso unos principios a tener en cuenta en la obtención de autopoliploides económicamente rentables:
- La especie debe tener un número cromosómico bajo.
- Su reproducción debe ser del tipo alógama.
- Su aprovechamiento económico se basa en las partes vegetativas.
La experiencia ha demostrado que la obtención de individuos con grados de ploidía superior al tetraploide no resultan de interés práctico, ya que no pueden competir con las formas diploides comerciales.
El centeno (Secale cereale) es la única especie en la que la autopoliploidía ha resultado práctica en lo referente al aprovechamiento de la semilla. En Japón, Inglaterra y los Estados Unidos se cultivan variedades tetraploides de uva y en Japón y los Estados Unidos se ha comercializado sandías triploides, ya que estas no presentan semillas. En lo referente al aprovechamiento de la parte vegetativa, en Suecia se han comercializado formas tetraploides de tréboles del género Trifolium sp. y forrajeras del género Brassica sp. El triploide de la remolacha azucarera es empleado en todo el mundo. La autopoliploidía también puede ofrecer buenas perspectivas en la mejora de plantas ornamentales.
Uso de alopoliploides
Cuando se hibridan especies vegetales, a veces el híbrido interespecífico resulta presentar una esterilidad gamética cromosómica, es decir, irregularidades meióticas debidas en muchos casos a la falta de afinidad entre los genomas paternos. El uso de la colchicina permite la obtención de híbridos con dos cromosomas homólogos para cada tipo cromosómico de cada genoma parental, resolviéndose el problema de la esterilidad gamética cromosómica, admitiéndose una relación entre la regularidad meiótica y la fertilidad. Cuanto más divergentes sean los genomas parentales, menor número de quiasmas se darán entre cromosomas homeólogos, por lo que la meiosis será más regular y la fertilidad mayor.
Entre los casos de alopoliploides inducidos, cabe destacar Triticale, un cereal doméstico obtenido a partir del trigo (Triticum aestivum) y del centeno (Secale cereale).
Referencias
- Marques, I.; Loureiro, J.; Draper, D.; Castro, M.; Castro, S. (1 de enero de 2018). «How much do we know about the frequency of hybridisation and polyploidy in the Mediterranean region?». Plant Biology (en inglés) 20: 21-37. ISSN 1438-8677. doi:10.1111/plb.12639. Consultado el 13 de marzo de 2018.
- Müntzing, A. 1936. The evolutionary significance of autopolyploidy. Hereditas 21: 263–378
- Darlington, C. D. 1937. Recent advances in cytology, 2nd ed. P. Blakiston’s, Philadelphia, Pennsylvania, USA.
- Stebbins, G. L. 1950. Variation and evolution in plants. Columbia University Press, New York, New York, USA.
- Grant, V. 1981. Plant speciation, 2nd ed. Columbia University Press, New York, New York, USA.
- Goldblatt, P. 1980. Polyploidy in angiosperms: Monocotyledons. In W. H. Lewis [ed.], Polyploidy: Biological relevance, 219–239. Plenum Press, New York, New York, USA.
- BOHLEN J, RITTERBUSCH D. Which factors affect sex ratio of spined loach (genus Cobitis) in Lake Müggelsee. Environmental Biology of Fishes 2000; 59:347-52
- KLINKHARDT M, TESCHE M, GREVEN H. Database of fish chromosomes. Magdeburg: Westarp Wissenschaften, 1995
- REDDY PVGK. Genetic resources of Indian major carps. FAO Fisheries Technical Paper 387. Rome: FAO, 1999
- GRIFFITH OL, MOODIE GEE, CIVETTA A. Genome size and longevity in fish. Experimental Gerontology 2003; 38:333-7
- Nakamura, H. 1985. A review of molluscan cytogenetic information based on the CISMOCH-Computerized Index System for Molluscan Chromosomes. Bivalvia, Polyplacophora and Cephalopoda. Venus 44: 193-225.
- Lee, T. 1999. Polyploidy and meiosis in the freshwater clam Sphaerium striatinum (Lamarck) and chromosome numbers in the Sphaeriidae (Bivalvia, Veneroida). Cytologia 64: 247-252.
- Lee, T. & D. Ó Foighil. 2002. 6-Phosphogluconate deshydrogenase (PGD) allele phylogeny is incongruent with a recent origin of polyploidization in some North American Sphaeriidae (Mollusca, Bivalvia). Molecular Phylogenetics and Evolution 25: 112-124.
- Texto de referencia
- Ramsey, J. (January 1998). «Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants». Annual Review of Ecology and Systematics 29 (1): 467-501. doi:10.1146/annurev.ecolsys.29.1.467.
- Schmid, M.; Evans, B. J.; Bogart, J. P. (2015). «Polyploidy in Amphibia». Cytogenetic and Genome Research 145 (3–4): 315-330. PMID 26112701. doi:10.1159/000431388.
- Janik, J.; Stevenson, E.C. 1955. The effects of polyploidy on sex expression in spinach. J. Hered., 46: 151-156.
Bibliografía
- Lacadena, J.R. 1996. Citogenética. Editorial Complutense, Madrid. ISBN 84-89365-58-X
- soltis
- (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
 Datos: Q213410
Datos: Q213410