Tomistominae
Tomistominae es una subfamilia de crocodilianos que incluye a una especie viviente, el falso gavial. Muchas más especies hoy extinguidas se conocen, extendiendo el rango temporal de la subfamilia hasta el período Eoceno. En contraste con el falso gavial, que es una especie de agua dulce que vive solo en el sudeste de Asia, los tomistominos extintos tenían una distribución global y vivían en estuarios y a lo largo de las costas.
| Tomistominae | ||
|---|---|---|
| Rango temporal: 55 Ma - 0 Ma Eoceno - Reciente | ||
.jpg.webp) Falso gavial, Tomistoma schlegelii | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Reptilia | |
| (sin rango): | Archosauria | |
| Orden: | Crocodylia | |
| Superfamilia: | Crocodyloidea / Gavialoidea | |
| Familia: | Crocodylidae / Gavialidae | |
| Subfamilia: |
Tomistominae Kälin, 1955 | |
| Géneros | ||
| ||
La clasificación de los tomistominos dentro de Crocodylia es incierta; tradicionalmente se los ha considerado parte de Crocodyloidea, pero las evidencias moleculares sugieren que están más relacionados con los verdaderos gaviales, es decir, miembros de la superfamilia Gavialoidea.[1]
Descripción
Los tomistominos tienen hocicos largos y estrechos, similares a los de los gaviales. El actual falso gavial vive en agua dulce y usa su hocico estrecho y sus dientes agudos para atrapar peces, si bien los verdaderos gaviales están más adaptados a la dieta piscívora, o consumo de peces. A pesar de las similitud con los gaviales, la forma de los huesos en los cráneos de los tomistominos los vinculan con los cocodrilos propiamente dichos. Por ejemplo, tanto los tomistominos como los cocodrilos tienen barras postorbitales delgadas detrás de las cuencas oculares y un gran alvéolo para el quinto diente maxilar. El hueso esplenial de la mandíbula es largo y estrecho, formando una distintiva forma de "V" no vista en los gaviales.
Historia evolutiva
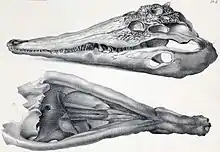
Los tomistominos aparecieron por primera vez en Europa y el África del Norte. El más antiguo tomistomino conocido es Kentisuchus spenceri de Inglaterra, aunque un posible fósil de tomistomino del Paleoceno de España es aún más antiguo.[2] Otros tomistominos tempranos incluyen a Maroccosuchus zennaroi de Marruecos y Dollosuchus dixoni de Bélgica. Estos primeros tomistominos habitaron el mar de Tetis, que cubría la mayor parte de Europa y el Norte de África durante el Paleógeno. Varios tomistominos primitivos han sido hallados en depósitos de costas marinas, sugiriendo que vivieron en las costas o en estuarios. Los gavialoides extintos parecen haber sido animales también costeros. El estilo de vida marino de estas primeras formas probablemente permitió a los tomistominos expandirse por el Tetis, formando una población en el norte en Europa y una sureña en África.[2]
Más tarde en el Eoceno y el Oligoceno, los tomistominos se expanden por Asia. Las especies de mediados del Eoceno Ferganosuchus planus y Dollosuchus zajsanicus son conocidas de Kazajistán y Kirguizistán. Los tomistominos alcanzaron China y Taiwán con las especies del Eoceno tardío "Tomistoma" petrolicum y Penghusuchus pani del Mioceno.[3] Otra especie, "Tomistoma" tandoni, vivió en la India durante el Eoceno medio. En este tiempo, el subcontinente Indio estaba separado del continente asiático, creando una barrera para las especies que no toleraban el agua salada. El mar de Obik, que separaba Europa de Asia, también impedía el cruce. Los tomistominos fueron capaces de cruzar estas áreas, lo que indica que eran capaces de tolerar el agua salada.[2]
Los tomistominos cruzaron el océano Atlántico y se expandieron por las Américas en el Oligoceno, el Mioceno y el Plioceno. El más antiguo de los tomistominos neotropicales conocido es Charactosuchus kuleri de Jamaica. Una cercana relación se ha propuesto entre C. kuleri y D. zajsanicus de Bélgica, sugiriendo que los tomistominos cruzaron de Europa a América a través del puente terrestre De Geer que conectaba a Noruega con Groenlandia y a Norteamérica o el puente terrestre de Thule que conectaba a Escocia, Islandia y Groenlandia con Norteamérica. El género Thecachampsa estaba presente a lo largo de la costa este de Norteamérica desde el Oligoceno hasta el Plioceno.[2]

Los tomistominos desaparecieron de Europa durante el Oligoceno pero retornaron a fines de ese período. Se diversificaron y se hicieron comunes a mediados del Mioceno. Una especie, Tomistoma coppensi, es conocida del final del Mioceno de Uganda. La aparición de tomistominos en el África central es inusual debido a que hay poca evidencia de especies del Mioceno en el Norte de África, un área desde la cual debieron haber llegado tras atravesar Europa.[2]
Los tomistominos pueden haber viajado de África hacia Asia cuando Arabia colisionó con el continente eurasiático a principios del Mioceno. Sin embargo, los tomistominos asiáticos del Mioceno pueden ser descendientes de las especies que estaban presentes en el oriente asiático desde el Eoceno. Los tomistominos se expandieron en el subcontinente indio en este tiempo. Una especie, Rhamphosuchus crassidens fue uno de los mayores crocodilianos que hayan vivido, llegando a una longitud estimada de entre 8 a 11 metros. Nuevas especies como Toyotamaphimeia machikanensis estaban presentes en Japón en el Pleistoceno. En el sureste asiático sin embargo, hay poca evidencia fósil de los tomistominos que precedieron al falso gavial. Por lo tanto, su relación con las especies extintas no es clara.[2]
Filogenia
| Hipótesis de la filogenia de los tomistominos | ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
El falso gavial Tomistoma es similar al gavial Gavialis es que es longirrostrino, es decir, poseen hocicos largos y estrechos. Aunque los cocodrilos son también longirrostrinos, sus hocicos no son tan estrechos como los de Tomistoma y Gavialis. Detalles de la morfología de Tomistoma y Gavialis sugieren que apenas son parientes lejanos y que los hocicos largos evolucionaron independientemente en cada linaje como resultado de una evolución convergente. Morfológicamente, los tomistominos parecen estar más relacionados con los crocodílidos, compartiendo un más reciente ancestro común con los cocodrilos verdaderos (crocodylinae) que con los gaviales. Bajo esta filogenia, los gaviales divergieron de los ancestros comunes de los tomistominos, cocodrilos y caimanes hace unos 65 millones de años, en el Cretácico Superior.[4] Abajo un cladograma de Piras et al. (2007) colocando a los tomistominos dentro Crocodyloidea:[2]
| Crocodylia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Abajo está un cladograma de Shan et al. (2009):[3]
| Crocodyloidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Recientes análisis moleculares de los crocodilianos encontraron una cercana relación entre Tomistoma y Gavialis. De acuerdo a esta filogenia, las dos formas son taxones hermanos, más cercanamente relacionados el uno al otro que a cualquier otro tipo de crocodiliano viviente. Si es así, la divergencia de ambas formas debió ocurrir entre el Eoceno hasta mediados del Mioceno.[4] Si se considera que Tomistoma y Gavialis son parientes cercanos, las similitudes morfológicas entre Tomistoma y los cocodrilos serían ahora el caso de convergencia. En este caso, la convergencia lleva a un problema conocido como atracción de ramas largas, donde características similares, que evolucionaron de forma independiente en Tomistoma y los cocodrilos erróneamente hacen suponer que estos dos muy derivados grupos están estrechamente relacionados. En efecto, estas características compartidas ocultarían la cercana relación entre Tomistoma y Gavialis.[5]
La atracción de ramas largas puede ser resuelta si las diferencias son observadas en los miembros ancestrales de cada grupo. Si los dos grupos están cercanamente relacionados, los miembros tempranos podrían tener rasgos primitivos del ancestro común y deberían carecer de rasgos derivados que causan la atracción de ramas largas. Los fósiles de tomistominos no resuelven la atracción de ramas largas cuando sus características son añadidas en los análisis filogenéticos, entonces aún permanece el conflicto entre las filogenias moleculares y morfológicas.[5]
Algunos estudios han combinado características morfológicas y moleculares en sus análisis. Los análisis que incluyen características morfológicas pero siguen usando bastante las secuencias de ADN encuentran una relación cercana entre Tomistoma y Gavialis. Los análisis que usan más características de los huesos y fósiles sostienen la cercana relación de Tomistoma con los cocodrilos.[5]
Referencias
- Gatesy, Jorge; Amato, G.; Norell, M.; DeSalle, R.; Hayashi, C. (2003). «Combined support for wholesale taxic atavism in gavialine crocodylians». Systematic Biology 52 (3): 403-422. doi:10.1080/1063515035019703. Archivado desde el original el 22 de junio de 2019. Consultado el 17 de julio de 2011.
- Piras, P.; Delfino, M.; Del Favero, L.; Kotsakis, T. (2007). «Phylogenetic position of the crocodylian Megadontosuchus arduini and tomistomine palaeobiogeography». Acta Palaeontologica Polonica 52 (2): 315-328.
- Shan, Hsi-yin; Wu, Xiao-chun; Cheng, Yen-nien; Sato, Tamaki (2009). «A new tomistomine (Crocodylia) from the Miocene of Taiwan». Canadian Journal of Earth Sciences 46 (7): 529-555. doi:10.1139/E09-036.
- Piras, P.; Colangelo, P.; Adams, D.C.; Buscalioni, A.; Cubo, J.; Kotsakis, T.; Meloro, C.; Raia, P. (2010). «The Gavialis-Tomistoma debate: the contribution of skull ontogenetic allometry and growth trajectories to the study of crocodylian relationships». Evolution & Development 12 (6): 568-579. doi:10.1111/j.1525-142X.2010.00442.x.
- Brochu, C. A. (2003). «Phylogenetic approaches toward crocodylian history». Annual Review of Earth and Planetary Sciences 31: 357-97. doi:10.1146/annurev.earth.31.100901.141308. Archivado desde el original el 2 de abril de 2015. Consultado el 17 de julio de 2011.
 Portal:Saurópsidos. Contenido relacionado con Saurópsidos.
Portal:Saurópsidos. Contenido relacionado con Saurópsidos.
| Control de autoridades |
|
|---|
 Datos: Q2448703
Datos: Q2448703 Multimedia: Tomistominae / Q2448703
Multimedia: Tomistominae / Q2448703 Especies: Tomistominae
Especies: Tomistominae