Poliovirus
El poliovirus es un virus perteneciente al género de enterovirus, es de la familia Picornaviridae. Es el agente causante de la poliomielitis en humanos.[1] Es un pequeño virus ARN (ácido ribonucleico), de cerca de 300 Angstrom de diámetro, cuyo genoma está compuesto por una hélice simple de ARN en sentido positivo,(virus monocatenario positivo); contiene alrededor de 7500 bases de longitud.[2] El poliovirus fue aislado en 1908 por Karl Landsteiner y Erwin Popper. En 1981, el genoma del poliovirus fue publicado por dos diferentes grupos de investigadores, uno del MIT y el otro de la Universidad de Nueva York.[3] Debido a su corto genoma y su simple composición—solamente una molécula de ARN y una cubierta proteica icosaédrica sin envoltura que la cubre— el poliovirus está considerado como uno de los virus más simples.[4]
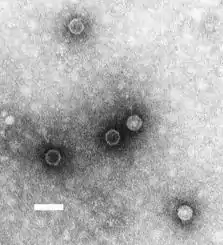

Poliovirus en la humanidad
Los poliovirus, aparentemente, han sido patógenos importantes de los humanos durante mucho tiempo. Existen representaciones en estelas egipcias que datan de hace más de 3500 años sugieren que el poliovirus estuvo presente en el Egipto antiguo. Aunque el virus ha estado muy distribuido en humanos durante miles de años, no hay evidencias de poliomielitis en poblaciones humanas hasta hace 200 años donde aparentemente el virus se extendió mundialmente. Encuestas en los Estados Unidos en los años 30 y 40, indicaban que alrededor del 80-100% de adultos habían sido infectados por poliovirus en algún momento de sus vidas.
Aunque el poliovirus estaba muy extendido, las epidemias de poliomielitis de grandes proporciones evolucionaron solo hasta el siglo XX, fenómeno que se explica como resultado de cambios en el comportamiento de la humanidad. Originalmente, el virus altamente infeccioso era contraído por infantes poco después de nacidos, cuando aún estaban protegidos por anticuerpos maternales. Esta infección natural servía para inmunizar al infante, protegiéndolo de por vida. Sin embargo, cuando la cadena de inmunización fue interrumpida por la eliminación del virus del ambiente por el desarrollo de condiciones higiénicas y niños crecían desprotegidos, dando lugar a poblaciones susceptibles. Si el virus invadía dichas poblaciones, las epidemias evolucionaban rápidamente.[5]
Ciclo de vida
El poliovirus infecta células humanas por unión con un receptor similar al de la inmunoglobulina CD155 (también conocido como receptor del poliovirus (PVR)) en la superficie de la célula humana.[6] La interacción del poliovirus y el CD155 facilita un cambio conformacional irreversible de la partícula viral necesario para la entrada viral.[7][8] El mecanismo preciso del poliovirus para entrar a la célula huésped no está firmemente dirimido.[9] Unido a la membrana celular, la entrada del ácido nucleico viral se cree que puede ocurrir de 2 formas: mediante la formación de un poro en la membrana a través de la cual el ARN viral es “inyectado” dentro del citoplasma, o bien el virus es ingresado vía endocitosis inducida por el receptor.[10] Por cualquiera de esos mecanismos, la estrategia de ingreso del poliovirus es muy ineficiente; solo está disponible para iniciar una infección cerca del 1 % del tiempo.[11]

El poliovirus es un virus ARN monocatenario positivo. El genoma encerrado en la partícula vírica puede ser usado como ARN mensajero e inmediatamente traducido por la célula huésped. Al entrar el virus, secuestra la maquinaria de traducción celular, causando la inhibición de la síntesis de proteínas celulares en favor de la producción de proteínas específicas del virus. A diferencia del ARN mensajero celular, el extremo 5' del ARN del poliovirus es extremadamente largo, en torno a 700 nucleótidos, y es altamente estructurado. Esta región del genoma es llamada sitio interno de entrada al ribosoma (IRES) y dirige la traducción del ARN viral. Las mutaciones genéticas en esta área impiden la producción de proteínas virales.[12][13][14]
El ARN mensajero es traducido a un largo polipéptido. Este polipéptido es auto-cortado por proteasas internas en aproximadamente 10 proteínas virales individuales, entre las que se incluyen:[4][11]
- 3Dpol, una ARN dependiente de ARN polimerasa, que permite al ARN genómico del polio ser copiado dentro de la célula huésped.

- 2Apro y 3Cpro/3CDpro, proteasas que parten el polipétido viral
- VPg (3B), una pequeña proteína que se funde con el ARN viral y es necesaria para la síntesis de hebras de ARN virales positivas y negativas
- 2BC, 2B, 2C, 3AB, 3A, 3B proteínas que componen el complejo de proteínas necesario para la replicación viral
- VP0, VP1, VP2, VP3, VP4 proteínas del cápside viral
El ensamblaje de nuevas partículas de virus, ( i.e. el empaquetado de genomas de progenie en cápsides que pueden sobrevivir fuera de la célula hospedante) está pobremente entendido.[10] El poliovirus completamente ensamblado abandona los confines de su hospedante tras las 4-6 horas siguientes al inicio de la infección.[16] El mecanismo de salida de los virus de la célula es poco claro,[2] pero cada una de ellas antes de morir puede lanzar más de 10 000 viriones de polio.[16]
Patogénesis
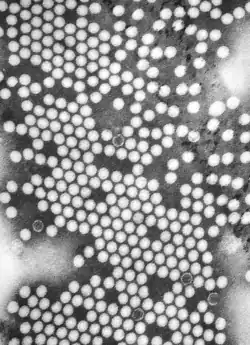
La infección por el poliovirus está limitada por la expresión del receptor CD155, que se encuentra solo en las células de los humanos, primates superiores, y el mono del Viejo Mundo. El poliovirus es, sin embargo, estrictamente un patógeno humano y no infecta de forma natural ninguna otra especie.[17]
Típicamente, la infección por el poliovirus conlleva la replicación del virus dentro del tracto gastrointestinal y la ulterior excreción del virus en las heces. En el 95% de los casos solo se produce la presencia primaria, transitoria del virus en la corriente sanguínea (llamada viremia) y la infección por el poliovirus es asintomática. En cerca del 5% de los casos, el virus se disemina y replica en otros sitios como grasa parda, sistema retículo endotelial y músculo. Esta replicación causa viremia secundaria y conduce al desarrollo de síntomas menores como fiebre, cefalea, y dolor de garganta.[18]
En cerca de 1-2% de infecciones por poliovirus el virus entra en el sistema nervioso central y se replica en la motoneurona ubicada en la médula espinal, tallo cerebral o corteza motora, resultando destrucción selectiva de neuronas motoras, que causa parálisis temporal o permanente y, en casos raros, paro respiratorio y muerte. En muchos aspectos la fase neurológica de la infección puede considerarse como desviación accidental de la infección gastrointestinal. Los mecanismos por los cuales el poliovirus se disemina al sistema nervioso central son poco comprendidos,[10] sin embargo se han sugerido dos teorías para explicar la desviación del virus.
La primera hipótesis es que el virus pasa directamente de la sangre al sistema nervioso central cruzando la barrera hematoencefalica, sin unirse a su receptor celular CD 155.[19] La segunda hipótesis sugiere que el virus es transportado del músculo a la médula espinal a través de transporte axonal retrogrado.[20] Ambas teorías requieren que el virus este presente en la sangre, y se ha demostrado que el poliovirus se puede unir y replicar en monocitos en la sangre,[21] quienes pueden entonces estar involucrados en la diseminación del virus de los sitios primarios de infección (el intestino) a la circulación y potencialmente el sistema nervioso central.
Evasión al sistema inmunitario
El Poliovirus utiliza dos mecanismos claves para evadir el sistema inmunitario. Primero, es capaz de sobrevivir en las condiciones de pH altamente ácido del tracto gastrointestinal, permitiendo al virus infectar al hospedador y diseminarse por el sistema linfático.[4] Segundo, porque puede replicarse muy rápidamente - el virus abruma los órganos del hospedador antes de que se produzca una respuesta inmune.[22] En individuos inmnocompetentes, anticuerpos contra el poliovirus están presentes en las amígdalas, y tracto gastrointestinal, (específicamente IgA) y son capaces de bloquear la replicación del poliovirus; anticuerpos IgG e IgM (séricos) contra poliovirus pueden prevenir la diseminación del virus a las neuronas motoras del sistema nervioso central.[16]
Serotipos
Existen tres serotipos de poliovirus, PV1 (Mahoney), PV2 (Lansing), y PV3 (León), cada uno con cápsides proteicas diferentes. El PV1 es la forma más común encontrada en la naturaleza, sin embargo las tres formas son muy infecciosas. Los tres tipos de poliovirus son estructuralmente similares a otros enterovirus humanos, coxsackievirus, echovirus, y pararhinovirus humanos, que también utilizan imitadores de inmunoglobulinas para entrar en las células huésped. La infección con un tipo de poliovirus no proporciona inmunidad contra los otros tipos, sin embargo segundos ataques dentro de la misma persona son extremadamente raros.
Referencias
- Ryan KJ; Ray CG (editors) (2004). Sherris Medical Microbiology (4th ed. edición). McGraw Hill. ISBN 0-8385-8529-9.
- Hogle J. «Poliovirus cell entry: common structural themes in viral cell entry pathways». Annu Rev Microbiol 56: 677-702. PMID 12142481.
- Kitamura N, Semler B, Rothberg P, Larsen G, Adler C, Dorner A, Emini E, Hanecak R, Lee J, van der Werf S, Anderson C, Wimmer E (1981). «Primary structure, gene organization and polypeptide expression of poliovirus RNA». Nature 291 (5816): 547-53. PMID 6264310.
- David S. Goodsell (1998). The machinery of life. Nueva York: Copernicus. ISBN 0-387-98273-6.
- Strauss, James (2008). «3». Viruses and Human Disease (en inglés). ELSEVIER. ISBN 978-0-12-373741-0.
- He Y, Mueller S, Chipman P, Bator C, Peng X, Bowman V, Mukhopadhyay S, Wimmer E, Kuhn R, Rossmann M (2003). «Complexes of poliovirus serotypes with their common cellular receptor, CD155». J Virol 77 (8): 4827-35. PMID 12663789.
- Kaplan G, Freistadt MS, Racaniello VR (1 de octubre de 1990). «Neutralization of poliovirus by cell receptors expressed in insect cells». J. Virol. 64 (10): 4697-702. PMC 247955. PMID 2168959.
- Gomez Yafal A, Kaplan G, Racaniello VR, Hogle, JM (1993). «Characterization of poliovirus conformational alteration mediated by soluble cell receptors». Virology 197 (1): 501-505. PMID 8212594. doi:10.1006/viro.1993.1621.
- (via NCBI Bookshelf) ISBN 0-9631172-1-1 Picornaviruses: The Enteroviruses: Polioviruses in: Baron's Medical Microbiology (Baron S et al, eds.)] (4th ed. edición). Univ of Texas Medical Branch. 1996.
- Mueller S, Wimmer E, Cello J (2005). «Poliovirus and poliomyelitis: a tale of guts, brains, and an accidental event». Virus Res 111 (2): 175-93. PMID 15885840.
- Charles Chan and Roberto Neisa. "Poliomyelitis". Brown University.
- Chen CY, Sarnow P (1995). «Initiation of protein synthesis by the eukaryotic translational apparatus on circular RNAs». Science 268 (5209): 415-7. PMID 7536344. doi:10.1126/science.7536344.
- Pelletier J, Sonenberg N (1988). «Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA». Nature 334 (6180): 320-325. PMID 2839775. doi:10.1038/334320a0.
- Jang SK, Kräusslich HG, Nicklin MJ, Duke GM, Palmenberg AC, Wimmer E (1 de agosto de 1988). «A segment of the 5' nontranslated region of encephalomyocarditis virus RNA directs internal entry of ribosomes during in vitro translation». J. Virol. 62 (8): 2636-43. PMC 253694. PMID 2839690.
- De Jesus NH (2007). «Epidemics to eradication: the modern history of poliomyelitis». Virol. J. 4 (1): 70. PMC 1947962. PMID 17623069. doi:10.1186/1743-422X-4-70.
- Kew O, Sutter R, de Gourville E, Dowdle W, Pallansch M (2005). «Vaccine-derived polioviruses and the endgame strategy for global polio eradication». Annu Rev Microbiol 59: 587-635. PMID 16153180.
- Mueller S, Wimmer E (2003). «Recruitment of nectin-3 to cell-cell junctions through trans-heterophilic interaction with CD155, a vitronectin and poliovirus receptor that localizes to alpha(v)beta3 integrin-containing membrane microdomains». J Biol Chem 278 (33): 31251-60. PMID 12759359. Archivado desde el original el 18 de marzo de 2009. Consultado el 29 de mayo de 2007.
- SABIN A (1956). «Pathogenesis of poliomyelitis; reappraisal in the light of new data». Science 123 (3209): 1151-7. PMID 13337331.
- Yang W, Terasaki T, Shiroki K, Ohka S, Aoki J, Tanabe S, Nomura T, Terada E, Sugiyama Y, Nomoto A (1997). «Efficient delivery of circulating poliovirus to the central nervous system independently of poliovirus receptor». Virology 229 (2): 421-8. PMID 9126254.
- Ren R, Racaniello V (1992). «Poliovirus spreads from muscle to the central nervous system by neural pathways». J Infect Dis 166 (4): 747-52. PMID 1326581.
- Freistadt M, Eberle K (2000). «Hematopoietic cells from CD155-transgenic mice express CD155 and support poliovirus replication ex vivo». Microb Pathog 29 (4): 203-12. PMID 10993739.
- Racaniello V (2006). «One hundred years of poliovirus pathogenesis». Virology 344 (1): 9-16. PMID 16364730.
| Control de autoridades |
|
|---|
 Datos: Q12438936
Datos: Q12438936 Multimedia: Poliovirus / Q12438936
Multimedia: Poliovirus / Q12438936 Especies: Poliovirus
Especies: Poliovirus