Élément génétique égoïste
Les éléments génétiques égoïstes (SGEs pour Selfish genetic elements) sont des séquences d'ADN codantes ou non codantes pouvant tout de même s'étendre à des micro-organismes ou des organites favorisant leur propre transmission au détriment du reste du génome de l'organisme. Leurs effets sur l’organisme hôte (porteur du SGE) sont le plus souvent neutres voire nuisibles.
Il existe de nombreux éléments génétiques dits égoïstes ayant divers mécanismes favorisant leur propre transmission : des éléments transposables (TE), des éléments induisant une conversion génique biaisée ou des biais méiotiques (meiotic drive), des distorteurs de post-ségrégation mais aussi des micro-organismes et organites héritables[1].
Historique
Les SGEs ont été sujets à une longue controverse historique.
En 1947, Ostergren indique que les chromosomes surnuméraires (aussi appelés chromosomes B) possèdent un mécanisme induisant un biais dans la méiose. Les chromosomes B semblaient donc se comporter comme des parasites génétiques c’est ainsi que pour la première fois un élément génétique fut considéré comme parasitaire.
En 1976, Dawkins introduit le terme de « gène égoïste » dans son livre intitulé The Selfish Gene. Il considérait tous les gènes comme étant égoïstes, dans le sens où ils sont sélectionnés pour produire un phénotype qui augmenterait leur fréquence dans les futures générations.
En 1988, bien que ces concepts soient encore contraires à l’opinion de la majorité des biologistes moléculaires, émerge le modèle des éléments génétiques égoïstes ou SGE. Ce modèle décrit la possibilité d'interactions entre éléments génétiques d'un même génome : coopérative (mutualisme), neutre (commensal) ou encore égoïste (parasitaire). De ces interactions peuvent alors découler des conflits génétiques lorsque les éléments en question diffèrent dans leur transmission. Lorsque la transmission de certains éléments se fait au détriment d'une partie du génome ou de l'individu lui-même, on parle alors « d’élément génétique égoïste ».
Pour la suite, c'est le modèle de 1988 qui sera retenu.
Parmi ces conflits génétiques, on peut distinguer des conflits intra-génomiques (entre éléments du même génome) tels que les biais méiotiques ou les transposons et des conflits inter-génomiques (entre génomes différents) tels que les organites et autres micro-organismes.
Aujourd'hui encore, de nombreux éléments génétiques sont découverts comme pouvant être égoïstes et cela dans les taxons les plus divers : des bactéries, aux animaux et végétaux[2],[1].
Les différents SGE
Transposons et autres éléments mobiles
Parmi ces éléments on peut citer : les plasmides, les virus endogènes et les éléments transposables (TE).
Encourageant son acquisition et son maintien dans la bactérie hôte, le plasmide est un exemple d'élément génétique mobile pouvant être décrit comme égoïste. Cet élément est connu pour pouvoir apporter des effets bénéfiques chez leur hôte. L'exemple le plus remarquable serait la résistance aux antibiotiques.
Les éléments transposables ont la capacité de se dupliquer et de se déplacer, ce qui peut mener à son accumulation dans le génome hôte. Leurs duplications et leurs insertions aléatoires peuvent en effet perturber des séquences codantes ou des régions de cis-régulations ; ils sont donc considérés comme des agents mutagènes. Ces effets néfastes s'ajoutent au coût énergétique qu'ils engendrent dans la cellule hôte lors de leur transcription et traduction. De ce fait, il y a une forte sélection dans le génome eucaryote pour favoriser l'évolution d'une machinerie supprimant l'activité des éléments transposables.
- Le mécanisme principal est la méthylation de l’ADN au niveau des résidus cytosines qui permet entre autres la répression des éléments transposables.
- Les ARN interférents (ARNi) induisent la non-expression de certains gènes et notamment des transposons. Ce sont des ARN messagers (ARNm) capables de complémenter une autre séquence d'ARNm, conduisant ainsi à une forme d'ARNm double brin qui sera ensuite dégradée. Ce mécanisme conduit ainsi à une diminution totale ou partielle de l'expression du gène codant l'ARNm complémentaire.
En réponse à cette machinerie de suppression mise en place chez la cellule, les TE développent leurs propres mécanismes pour se maintenir dans le génome de leur hôte selon le principe de « course à l'armement ».
Parmi ces mécanismes, les TE peuvent occuper des lieux « refuges » dans le génome. Ces refuges peuvent être une position dans le génome où l'hôte ne peut ni déplacer ni éliminer l'insert ou un lieu réduisant le coût de la fitness pour l'hôte.
Ainsi les éléments insérés à proximité des gènes codant des protéines sont moins éliminés parce que cela pourrait induire des conséquences nuisibles pour le gène adjacent. Dans un lieu refuge, les inserts peuvent donc persister plus longtemps même s'ils sont légèrement délétères. S'ils y sont maintenus suffisamment longtemps, certains de ces fragments de TE insérés dans l'ADN peuvent finalement évoluer en éléments de cis-régulation ou de régulations post-transcriptionnelles.
Étant donné l'abondance des TE – jusqu'à 40 % du génome humain et 90 % pour le génome du blé – il est inévitable que certains d’entre eux soient recrutés par le génome pour donner de nouvelles fonctions cellulaires. C'est une forme de « domestication », de « co-option » ou encore d'exaptation[3],[2],[1].
Conversion génique biaisée
La conversion génique biaisée correspond à un biais de la réparation de l’ADN conduisant à l’enrichissement des séquences en paire de nucléotides GC.
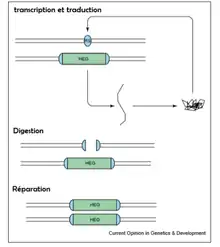
Les plus connus de ces mécanismes de conversions biaisées sont les gènes d'endonucléases homing (HEGs). Découverts dans des organites de fungi filamenteux, de plantes, d’algues et autres protistes, les HEGs sont peut-être l’un des plus simples des éléments génétiques égoïstes.
Chez les eucaryotes, bien que ne contribuant ni à la survie ni à la reproduction de l'espèce, ces éléments génétiques modifient leur taux de transmission d'une génération à l'autre, les proportions de Mendel s'en trouvant non respectées.
Chez les individus hétérozygotes, les HEGs ne sont présents que sur l'un des chromosomes homologues. Ces gènes codent des endonucléases – enzymes clivant l'ADN en un locus donné. Après synthèse, le chromosome porteur du gène étant protégé, l'enzyme va cliver uniquement la séquence de reconnaissance au niveau du chromosome homologue correspondant pour pouvoir y insérer le HEG précédemment dupliqué. Ce mécanisme, appelé « homing », permet donc la conversion d'une forme hétérozygote d'un gène à une forme homozygote, assurant ainsi sa transmission à la génération suivante.
Le succès de cette stratégie dépend des opportunités qu'ont les HEGs de se répandre, en d'autres termes, de la fréquence à laquelle un chromosome porteur des HEGs rencontre son homologue ne le portant pas. Cette fréquence varie beaucoup selon les taxons.
Une fois fixée dans la population, les pressions de sélection s’exerçant sur les HEGs disparaissent, ce qui peut entraîner leur dégénérescence. Il n’y aura donc plus de pression de sélection pour maintenir l'intégrité des HEGs qui seront alors plus vulnérables aux mutations pouvant perturber leur activité endonucléasique.
Une condition préalable est nécessaire aux HEGs pour qu'ils réussissent à se répandre : ils ne doivent pas causer trop de dommages à l'organisme hôte. Ceci peut avoir des conséquences sur l'évolution de leur séquence de spécificité (15-20pb) et de leur fonction de maturation.
Les HEGs peuvent aussi être domestiqués par leur hôte et sont couramment utilisés dans la recherche. En effet, ils sont utilisés pour faire de la cartographie de génome, pour étudier le système de réparation des chromosomes, etc. Ils seraient également un outil indispensable en ce qui concerne la thérapie génique humaine, ainsi que dans l'éradication d’espèces nuisibles[4],[5].
Biais méiotique
En général, la méiose est "juste". Les chromosomes homologues ont une probabilité égale de finir dans un gamète fonctionnel (ovocyte ou spermatozoïde). Il s'avère cependant que dans de nombreuses espèces, des chromosomes ou des régions chromosomiques particulières sont surreprésentés dans les gamètes transmis par rapport au reste du génome. Ce phénomène est ce que l'on appelle le biais méiotique (meiotic drive).
Ces biais méiotiques ont été mis en évidence chez de nombreuses espèces telles que les mouches, les souris, les moustiques, etc.
Certains systèmes ont été intensivement étudiés : les distorteurs de ségrégation (segregation distorter ou SD) chez Drosophila melanogaster, l'élément génétique sex-ratio (SR) chez D. pseudoobscura, le knob-containing chromosome chez le maïs, le locus T chez Mus musculus ainsi que le chromosome surnuméraire B présent chez de nombreux animaux et végétaux.
Ces biais méiotiques peuvent prendre 3 formes basiques :
- le vrai biais méiotique (« true » meiotic drive) comprenant par exemple les chromosomes B ou encore le centromere drive. Il s'agit d'une ségrégation préférentielle au pôle fonctionnel de l’ovocyte durant la gamétogenèse.
- la sur-réplication dans la lignée germinale du chromosome B, il est alors présent en un plus grand nombre de copies, augmentant alors sa transmission lors de la formation des gamètes.
- le « gamete killer » qui conduit à une élimination sélective ou une perturbation fonctionnelle du gamète qui ne le porte pas. Les mâles produisent un excès de gamètes qui vont donc entrer en compétition pour la fécondation, c'est donc chez eux que se retrouve le plus souvent cette forme de biais méiotique. Le gamete killer augmente ainsi la fécondation des ovocytes par les spermatozoïdes porteurs de l'élément génétique égoïste[1],[2].
« gamete killer », exemple du biais méiotique par sex-ratio (SR)
Chez D.pseudoobscura, les mâles porteurs d'un chromosome SR X (c'est-à-dire que le chromosome X du mâle porte l'élément génétique SR) transmettent uniquement le chromosome X à leur descendance. Par conséquent, les mâles porteurs de SR ne produisent que des femelles, et le nombre de mâles est corrélé négativement avec la fréquence de SR dans la population.
Bien que la méiose du mâle SR se déroule normalement, les spermatozoïdes ne portant pas l'élément génétique SR n'arriveront pas à maturité lors de la spermatogenèse. On peut alors facilement imaginer qu'un tel mécanisme conduirait à un remplacement de tous les chromosomes X réguliers et causer l'extinction de la population. Cependant, la fréquence actuelle de l'élément génétique SR montre une distribution clinale n'excédant pas les 20 % de la population.
Il existe des mécanismes limitant la diffusion de l'élément génétique SR :
- Un premier mécanisme semble être l'évolution de suppresseur génotypique ou de chromosome Y insensible.
- Par ailleurs, les populations SR ont une fitness négative : les femelles homozygotes possédant SR sont relativement stériles tandis que les mâles SR souffrent d'une diminution significative de leur fertilité, en partie due à la dégénérescence de leurs gamètes.
Mais ce qui peut particulièrement limiter la reproduction de leurs descendants et donc la transmission de l'élément SR est l’existence dans ces populations de la polyandrie. La polyandrie – fait qu'une femelle s'accouple avec plusieurs mâles – peut permettre de limiter la transmission des éléments SR. Elle régule la fréquence de SR en diminuant sa transmission, en augmentant la compétition entre les gamètes des différents mâles en concurrence. La polyandrie peut donc potentiellement prévenir l’augmentation de la fréquence de transmission ou l’élimination de l’élément SR dans la population. Le port de chromosomes SR X n'affecte cependant pas la tendance au ré-accouplement des femelles hôtes. Toutefois, chez les femelles sauvages, le taux de polyandrie peut augmenter en présence de mâles SR dans la population. Il semble exister une contre-adaptation chez certains mâles SR consistant à empêcher le ré-accouplement des femelles, la transmission de SR étant favorisée dans les populations mono-andrique. Dans ce cas, la compétition entre les gamètes des différents mâles est transférée à une compétition sexuelle entre les mâles.
Bien que la polyandrie, via les contre-adaptations des mâles SR, puisse augmenter les conflits entre individus et ainsi réprimer l'évolution de la sociabilité, elle permet de promouvoir l'harmonie dans le génome en supprimant l'augmentation des fréquences de transmission du SR[2],[6].
Chromosome B
Les chromosomes B sont des chromosomes supplémentaires qui ne sont pas essentiels pour la viabilité de l'organisme et ne modifient généralement pas leur phénotype. Ils sont souvent hétérochromatiques c’est-à-dire plus condensés que les chromosomes réguliers. Ils augmentent leur transmission durant la gamétogenèse par une sur-réplication dans les cellules germinales et/ou une ségrégation préférentielle dans le nucléus de l’ovocyte, plus que dans le globule polaire. Il existe différents mécanismes de transmission variant en fonction du type de chromosome B, de l’espèce hôte et des conditions environnementales. Ainsi le chromosome B n'est parfois présent que dans des populations en croissance dans un environnement favorable. C’est dans ces conditions que le chromosome B est moins dangereux et nuisible pour l'organisme qui le porte[2].
Distorteur de Post ségrégation (Post Segregation Distorters ou PSDs)
Après la fécondation, les PSDs diminuent la proportion des individus ne les portant pas. De manière générale, une modification arrive chez les parents. Cette modification doit être « sauvée » chez la descendance. Si l’élément PSD n'est pas transmis à la descendance, le sauvetage n'a pas lieu et des effets défavorables arrivent à la descendance. Le plus souvent ils agissent en tuant les individus qui n'ont pas reçu l'élément égoïste[3].
Plasmide tueur
Les plasmides tueurs sont présents chez de nombreuses bactéries et levures.
Ce plasmide code par exemple pour une « toxine » qui a la capacité de persister plus longtemps que la protéine « antidote » elle aussi codée par le plasmide tueur. Ainsi, si les cellules filles perdent le plasmide, la « toxine » étant plus persistante que l' « antidote », les cellules meurent. Cela empêche la perte du plasmide lors de la ségrégation ou le remplacement de ce plasmide par un compétiteur[3].
Le locus Medea
Le locus Medea (Maternal effect dominant embryonic arrest) chez les scarabées de farine Tribolium castaneum implique l'effet d'un allèle maternel qui tue la progéniture qui n’en a pas hérité. Même si les processus biochimiques ne sont toujours pas clairs, ils mettent probablement en jeu une modification de l’ovocyte (par des protéines ou de l'ARNm) qui ne peut être sauvé que par la présence du locus Medea chez le zygote[3].

L'incompatibilité cytoplasmique
C'est une incompatibilité entre l’ovocyte et le spermatozoïde due à une infection par une bactérie intracellulaire Wolbachia (qui est très répandue chez les insectes, arachnides, crustacés et nématodes). Des zygotes non infectés sont tués par des effets paternels dus aux pères infectés entraînant alors une nette augmentation de la fréquence d'individus infectés. Cette bactérie, qui agit en modifiant le spermatozoïde de l'hôte, va entraîner une incompatibilité létale avec le zygote si celui-ci ne présente pas la même souche de Wolbachia dans l’ovocyte pour le « sauver » de cette mutation. Une absence de ce « sauvetage » est donc incompatible et létale pour le zygote[3].
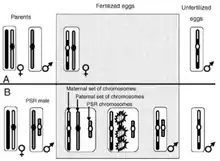
Le PSR (paternal sex-ratio)
Considéré comme le plus égoïste des SGE, le PSR est un chromosome B induisant une PSD. Il a été mis en évidence chez certains arthropodes haplodiploïdes. Il a été pour la première fois décrit chez la guêpe Nasonia vitripennis. Chez cette espèce, les femelles sont diploïdes et peuvent donner par parthénogenèse des mâles haploïdes ou, lors d'une fécondation par le mâle, des femelles diploïdes.
L'élément génétique PSR est un chromosome B surnuméraire porté par les mâles et transmis via les spermatozoïdes. Après fécondation, il cause la perte des chromosomes paternels, convertissant ainsi les ovocytes fécondés diploïdes – devant normalement se développer en femelle – en mâles haploïdes portant le PSR.
Le phénomène se présente ainsi : les mâles porteurs du PSR produisent des spermatozoïdes fonctionnels. Cependant, après fécondation, les chromosomes paternels – exception faite du chromosome B – forment une masse chromatinienne dense à la première division mitotique et sont perdus. Le zygote est alors « haploïsé » et se développe en un mâle fonctionnel qui porte le PSR. Le PSR est immunisé contre cette hyper-condensation.
Il semblerait cependant que les mâles PSR présentent une fitness égal ou réduit à celui des mâles sauvages. De ce fait, le PSR ne peut envahir la population, il faudrait pour cela que plus de 50 % des ovocytes soient fécondés. Le PSR pourrait cependant être utilisé pour le contrôle des espèces d'insectes haploïdes[7].
Organites et micro-organismes héritables
Les organites (comme les mitochondries ou les chloroplastes) peuvent avoir des intérêts génétiques qui divergent de ceux du génome nucléaire, ce qui mène, bien évidemment, à un conflit génétique. De manière générale, les intérêts des organites sont plus compréhensibles si on considère une cellule comme une population d'organites en compétition les uns par rapport aux autres pour se transmettre à la génération suivante.
Des micro-organismes peuvent avoir une relation mutualiste avec la cellule hôte qui les héberge mais cela ne les empêchent de pouvoir manipuler la reproduction de leur hôte d'une manière qui augmente leur propre transmission conduisant à une « reproduction parasitaire ». Leur transmission préférentielle passant par les femelles, ils participent à la conversion de mâles en femelles fonctionnelles, induisent des femelles par parthénogenèse, tuent les mâles et/ou forment des incompatibilités entre les deux types de gamètes[2].
Théorie mathématique
Une grande partie de la confusion concernant les idées sur les éléments génétiques égoïstes est centrée sur l'utilisation du langage et la façon dont les éléments et leur dynamique évolutive sont décrits[8]. Les modèles mathématiques permettent de donner les hypothèses et les règles a priori pour établir des déclarations mathématiques sur la dynamique attendue des éléments dans les populations. Les conséquences de la présence de tels éléments dans les génomes peuvent alors être explorées objectivement. Les mathématiques peuvent définir de manière très précise les différentes classes d'éléments en fonction de leur comportement au sein d'une population, en évitant tout verbiage distrayant sur les espoirs et les désirs intérieurs de gènes égoïstes avides. Il existe de nombreux bons exemples de cette approche, et cet article se concentre sur les distordeurs de ségrégation, les systèmes d'entraînement des gènes et les éléments transposables[8].
Distorsions de ségrégation
L'allèle t de la souris est un exemple classique de système de distorsion de ségrégation qui a été modélisé de manière très détaillée[9],[10]. Les hétérozygotes pour un haplotype t produisent plus de 90 % de leurs gamètes portant le t (voir Distorsion de ségrégation), et les homozygotes pour un haplotype t meurent à l'état d'embryon. Il peut en résulter un polymorphisme stable, dont la fréquence d'équilibre dépend de la force d'entraînement et de l'impact direct des haplotypes t sur la valeur adaptative. C'est un thème commun aux mathématiques des distorsions de ségrégation : pratiquement tous les exemples que nous connaissons impliquent un effet sélectif compensatoire, sans lequel l'allèle à transmission biaisée se fixerait et la distorsion de ségrégation ne se manifesterait plus. Lorsque les chromosomes sexuels subissent une distorsion de ségrégation, le rapport de masculinité de la population est modifié, ce qui rend ces systèmes particulièrement intéressants. Deux exemples classiques de distorsion de ségrégation impliquant les chromosomes sexuels sont les chromosomes X "Sex Ratio" de Drosophila pseudoobscura[11] et les suppresseurs d'entraînement du chromosome Y de Drosophila mediopunctata[12]. Un point crucial de la théorie des distorsions de ségrégation est que ce n'est pas parce qu'il existe des effets de fitness agissant contre le distorseur que cela garantit l'existence d'un polymorphisme stable. En fait, certains conducteurs de chromosomes sexuels peuvent produire une dynamique de fréquence avec des oscillations et des cycles sauvages[13].
Forçage génétique
L'idée de diffuser un gène dans une population comme moyen de contrôle de la population est en fait assez ancienne, et les modèles de la dynamique des chromosomes composés introduits remontent aux années 1970[14]. Par la suite, la théorie de la génétique des populations pour les endonucléases homing et les systèmes d'entraînement génique basés sur CRISPR est devenue beaucoup plus avancée[15],[16]. Un élément important de la modélisation de ces processus dans les populations naturelles est de considérer la réponse génétique dans la population cible. D'une part, toute population naturelle hébergera une variation génétique permanente, et cette variation pourrait bien inclure un polymorphisme dans les séquences homologues aux ARN guides, ou les bras d'homologie qui sont censés diriger la réparation. En outre, des hôtes différents et des constructions différentes peuvent avoir des taux très différents de jonction des extrémités non homologues, la forme de réparation qui aboutit à des allèles cassés ou résistants qui ne se propagent plus. La prise en compte complète des facteurs de l'hôte représente un défi considérable pour qu'une construction d'entraînement génique puisse atteindre la fixation, et Unckless et ses collègues[17] montrent qu'en fait les constructions actuelles sont assez loin de pouvoir atteindre des fréquences même modérées dans les populations naturelles. Il s'agit là d'un autre excellent exemple montrant que ce n'est pas parce qu'un élément semble avoir un fort avantage de transmission égoïste que sa propagation réussie peut dépendre de configurations subtiles d'autres paramètres dans la population[16].
Éléments transposables
Pour modéliser la dynamique des éléments transposables (TE) au sein d'un génome, il faut réaliser que les éléments se comportent comme une population au sein de chaque génome, et qu'ils peuvent sauter d'un génome haploïde à un autre par transfert horizontal. Les mathématiques doivent décrire les taux et les dépendances de ces événements de transfert. On a observé très tôt que le taux de saut de nombreux TE varie avec le nombre de copies, et les premiers modèles ont donc simplement utilisé une fonction empirique pour le taux de transposition. Cette méthode avait l'avantage de pouvoir être mesurée par des expériences en laboratoire, mais elle laissait ouverte la question de savoir pourquoi le taux diffère entre les éléments et avec le nombre de copies. Stan Sawyer et Daniel L. Hartl [18] ont adapté des modèles de ce type à une variété de TE bactériens, et ont obtenu des ajustements assez bons entre le nombre de copies et le taux de transmission et l'incidence des TE à l'échelle de la population. Les TE des organismes supérieurs, comme la drosophile, ont une dynamique très différente en raison du sexe, et Brian Charlesworth, Deborah Charlesworth, Charles Langley, John Brookfield et d'autres[19],[20],[21] ont modélisé l'évolution du nombre de copies de TE chez la drosophile et d'autres espèces. Ce qui est impressionnant dans tous ces efforts de modélisation, c'est la façon dont ils s'adaptaient aux données empiriques, étant donné que cela se passait des décennies avant la découverte du fait que la mouche hôte possède un puissant mécanisme de défense sous la forme de piRNAs. L'incorporation de la défense de l'hôte ainsi que de la dynamique des TE dans les modèles évolutifs de la régulation des TE n'en est encore qu'à ses débuts[22].
Hypothèses sur les conséquences évolutives des SGE
Les éléments génétiques égoïstes sont omniprésents dans le génome des eucaryotes et, dans beaucoup de cas, ils représentent même une grande partie de l'ADN de l'organisme. Rappelons par exemple que les transposons ont colonisé jusqu'à 90 % du génome du blé. Ces éléments génétiques égoïstes ont attiré l'attention des biologistes cherchant à expliquer les principaux changements ayant eu lieu au cours de l'évolution, ceci pour deux raisons : tout d'abord ils illustrent clairement l'existence d'une sélection au niveau du gène et de quelle manière cette sélection peut aller en faveur de certains éléments génétiques, même si ces derniers sont en défaveur de l'organisme ou de la population. De plus, ils peuvent promouvoir une évolution dans la structure génétique des espèces, voire mener à des spéciations.
Les hypothèses sur les conséquences des SGE sont nombreuses : ils seraient à l'origine de la différenciation mâle / femelle, de l'héritage uni-parental des organites, de la structure du génome, ainsi que de nombreux autres aspects phénotypiques, comme la diversification du déterminisme sexuel.
De plus en plus d’intérêts se portent sur les SGE et leur possible rôle dans l'isolation reproductive et la spéciation eucaryote. L'isolation reproductive correspond à des barrières qui vont empêcher totalement ou partiellement la reproduction entre deux populations. Elle peut se produire de différentes manières mais a toujours pour conséquence que peu voire aucun gène(s) sont échangé(s) entre les populations.
De récentes recherches ont montré que l'incompatibilité cytoplasmique due à la bactérie Wolbachia a un rôle sur l'accélération de l'isolement reproducteur, permettant ainsi à la divergence de continuer et à la spéciation de se faire. Généralement, l'incompatibilité est unidirectionnelle, c'est-à-dire qu'une seule des deux populations est infectée, ce qui n'est pas suffisant pour induire une isolation reproductive.
La bactérie peut néanmoins traverser les « frontières » entre ces populations et, en les infectant toutes les deux, éliminer l'incompatibilité cytoplasmique et rétablir les flux de gènes entre elles. Cependant, ce phénomène ne peut avoir lieu que si l’incompatibilité cytoplasmique unidirectionnelle est couplée à d'autres mécanismes (ex : mate discrimination).
Un autre exemple d'isolement reproductif entre deux populations implique cette fois-ci une incompatibilité cytoplasmique bidirectionnelle. On a dans ce cas une infection des deux population par des lignées différentes de Wolbachia, causant de l'incompatibilité dans les deux directions.
Wolbachia est ainsi l’un des principaux contributeurs à l'isolation reproductive dans certains systèmes mais ne fait pas encore l’unanimité auprès des chercheurs.
Toutefois, de récents travaux montrent que cette incompatibilité cytoplasmique (même incomplète) peut suffire à maintenir et permettre une divergence génétique entre des populations naturelles. Des recherches complémentaires sont toutefois nécessaires pour déterminer quelle est l'importance de cette bactérie dans la spéciation chez les Arthropodes et nématodes.
Dès 1967, Hamilton a reconnu que la fixation des chromosomes sexuels causant des biais méiotiques, pouvait conduire à une extinction de l'hôte. En effet, via une diminution de la proportion de mâles à cause d’un biais en faveur du chromosome X ou bien d’un manque de femelle à la suite d'un biais en faveur du chromosome Y, le SGE peut se répandre dans la population, ce qui peut, en principe, conduire à l’extinction de la dite population.
L'extinction n'est cependant pas une fatalité après l'invasion d'un distorteur de sex-ratio. Elle peut être évitée si la distorsion de sex-ratio est incomplète ou si l'élément ne conduit pas à une fixation dans la population. Cela profite à la production de femelles, car un seul mâle peut maintenir la fertilité de plusieurs femelles simultanément.
Les causes d'une extinction sont un sujet difficile de recherche, mais des preuves indirectes montrent que les distorsions du sex-ratio chez certaines souches peuvent se fixer et donc causer l'extinction de la population.
Bien qu'il soit compliqué de savoir si les éléments de distorteur du sex-ratio aient causé l'extinction de populations, trente ans de recherches ont permis de mettre en évidence que la fixation des chromosomes sexuels peut conduire à l'extinction de l’espèce hôte confirmant l’idée qu’en avait Hamilton.
Le problème est évidemment qu'il est plus facile d'étudier un système dans lequel les éléments égoïstes n'ont pas causé d'extinction plutôt que ceux dans lesquels ils l'ont fait[3].
Notes et références
- (en) JH Werren, Selfish genetic elements, genetic conflict, and evolutionary innovation., vol. 108, PNAS, , p. 10863-10870
- (en) JH Werren, U Nur et CI Wu, Selfish genetic elements., vol. 3, Trends in Ecology & Evolution, , p. 297-302
- (en) G Hurst et JH Werren, The role of selfish genetic elements in eukaryotic evolution., vol. 2, Nature Reviews Genetics, , p. 597-606
- (en) A Burt et V Koufopanou, Homing endonuclease genes : the rise and fall and rise again of a selfish element., vol. 14, Current Opinion in Genetics & Development, , p. 609-615
- (en) FW Doolittle et C Sapienza, Selfish genes, the penotype paradigm and genome evolution., vol. 284, Nature, , p. 601-603
- (en) A Tom, Does polyandry control population sex ratio via regulation of a selfish gene ?, vol. 281, Proceedings of the Royal Society B, , p. 1-8
- (en) JH Werren et R Stouthamer, PSR (paternal sex ratio) chromosomes: The ultimate selfish genetic elements., vol. 117, Genetica, , p. 85-101
- Gardner A, Welch JJ, « A formal theory of the selfish gene », Journal of Evolutionary Biology, vol. 24, no 8, , p. 1801–13 (PMID 21605218, DOI 10.1111/j.1420-9101.2011.02310.x, S2CID 14477476)
- Lyon MF, « Transmission ratio distortion in mice », Annual Review of Genetics, vol. 37, , p. 393–408 (PMID 14616067, DOI 10.1146/annurev.genet.37.110801.143030)
- Lewontin RC, Dunn LC, « The Evolutionary Dynamics of a Polymorphism in the House Mouse », Genetics, vol. 45, no 6, , p. 705–22 (PMID 17247957, PMCID 1210083, DOI 10.1093/genetics/45.6.705)
- Curtsinger JW, Feldman MW, « Experimental and Theoretical Analysis of the "Sex-Ratio" Polymorphism in Drosophila pseudoobscura », Genetics, vol. 94, no 2, , p. 445–66 (PMID 17249004, PMCID 1214151, DOI 10.1093/genetics/94.2.445)
- Carvalho AB, Vaz SC, Klaczko LB, « Polymorphism for Y-linked suppressors of sex-ratio in two natural populations of Drosophila mediopunctata », Genetics, vol. 146, no 3, , p. 891–902 (PMID 9215895, PMCID 1208059, DOI 10.1093/genetics/146.3.891)
- Clark AG, « Natural selection and Y-linked polymorphism », Genetics, vol. 115, no 3, , p. 569–77 (PMID 3569883, PMCID 1216358, DOI 10.1093/genetics/115.3.569)
- Fitz-Earle M, Holm DG, Suzuki DT, « Genetic control of insect population. I. Cage studies of chromosome replacement by compound autosomes in Drosophila melanogaster », Genetics, vol. 74, no 3, , p. 461–75 (PMID 4200686, PMCID 1212962, DOI 10.1093/genetics/74.3.461)
- Burt A, « Site-specific selfish genes as tools for the control and genetic engineering of natural populations », Proceedings. Biological Sciences, vol. 270, no 1518, , p. 921–8 (PMID 12803906, PMCID 1691325, DOI 10.1098/rspb.2002.2319)
- Deredec A, Burt A, Godfray HC, « The population genetics of using homing endonuclease genes in vector and pest management », Genetics, vol. 179, no 4, , p. 2013–26 (PMID 18660532, PMCID 2516076, DOI 10.1534/genetics.108.089037)
- Unckless RL, Clark AG, Messer PW, « Evolution of Resistance Against CRISPR/Cas9 Gene Drive », Genetics, vol. 205, no 2, , p. 827–841 (PMID 27941126, PMCID 5289854, DOI 10.1534/genetics.116.197285)
- Sawyer S, Hartl D, « Distribution of transposable elements in prokaryotes », Theoretical Population Biology, vol. 30, no 1, , p. 1–16 (PMID 3018953, DOI 10.1016/0040-5809(86)90021-3)
- Charlesworth B, Langley CH, « The evolution of self-regulated transposition of transposable elements », Genetics, vol. 112, no 2, , p. 359–83 (PMID 3000868, PMCID 1202706, DOI 10.1093/genetics/112.2.359)
- Brookfield JF, Badge RM, « Population genetics models of transposable elements », Genetica, vol. 100, nos 1–3, , p. 281–94 (PMID 9440281, DOI 10.1023/A:1018310418744, S2CID 40644313)
- Charlesworth B, Charlesworth D, « The population dynamics of transposable elements. », Genet. Res., vol. 42, , p. 1–27 (DOI 10.1017/S0016672300021455
 )
) - Lu J, Clark AG, « Population dynamics of PIWI-interacting RNAs (piRNAs) and their targets in Drosophila », Genome Research, vol. 20, no 2, , p. 212–27 (PMID 19948818, PMCID 2813477, DOI 10.1101/gr.095406.109)
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire  Portail origine et évolution du vivant
Portail origine et évolution du vivant