BMP4
La BMP4, soit la protéine morphogénétique osseuse 4 (BMP, de l'anglais bone morphogenetic protein), est une protéine appartenant à la superfamille des facteurs de croissance transformants bêta TGF-β (de l'anglais transforming growth factor beta), qui regroupe elle-même la famille des protéines morphogénétiques osseuses[1]. Les protéines de la famille des TGF-β affectent notamment la croissance et la différenciation cellulaire[2].
Le gène responsable de la production de BMP4 est situé sur le 14e chromosome, à la position cytogénétique 14q22.2[3]. Chez l'humain, ce gène contient 4 exons fonctionnels, donnant lors de la transcription 11 ARNm différents et un polypeptide de 408 acides aminés à la suite de la traduction[1],[3]. BMP4 est trouvée chez l'ensemble des vertébrés et est extrêmement conservée au cours de l'évolution[2]. Durant le développement embryonnaire, cette protéine possède notamment un rôle dans la formation du mésoderme, la formation des dents, le développement squelettique, la formation de la moelle osseuse, la morphogenèse des membres et l'induction de la plaque neurale[1].
Découverte
Les BMP ont été découvertes pour la première fois par Marshall R. Urist en 1965, à la suite d'un isolement de trois protéines provenant de la déminéralisation d'os bovins[4]. Lorsqu'injectées dans des animaux, ces protéines induisaient la croissance du tissu osseux, plus particulièrement la formation de cartilage ectopique et d'os endochondraux.
Voie de signalisation
Lors de la signalisation, les BMP se lient à des récepteurs spécifiques à la surface de la cellule. Ces récepteurs, appelés BMPR (pour BMP-receptor), sont des récepteurs kinases sérine/thréonine. Via un groupe de protéines spécialisées nommées Smads, le signal produit par les BMP subit une transduction par les BMPR. Ce signal affecte ensuite la régulation des gènes cibles en aval[1],[2].
Il s'agit d'un processus extrêmement complexe pouvant être régulé de plusieurs façons, dont notamment un groupe de protéines inhibitrices extracellulaires de la signalisation qui agissent en se liant aux ligands des BMP. Dans le cas de BMP4, ses principaux inhibiteurs sont Noggin et Chordin[1].
Du point de vue extracellulaire et dépendamment du type de cellule ciblée, de la concentration de BMP4 et du moment du signal, BMP4 peut provoquer la prolifération cellulaire, la différenciation cellulaire ou l'apoptose. Il a été suggéré qu'une forte concentration de BMP4 aurait tendance à provoquer la différenciation, tandis qu'une faible concentration résulterait en une prolifération[5].
Développement de la morphologie craniofaciale chez les vertébrés
La morphologie craniofaciale détermine de quoi peut se nourrir une espèce, ainsi que son efficacité d'alimentation. Il va de soi que lorsque des espèces adoptent une alimentation spécialisée, il y a également un changement de la forme craniofaciale et de la fonction qui y est associée.
Il a été déterminé que BMP4 et CaM (calmoduline) ont un rôle majeur dans la détermination de la morphologie du bec des oiseaux et de la mâchoire des vertébrés lors du développement embryonnaire[6],[7].
Détermination de la morphologie du bec chez Geospiza
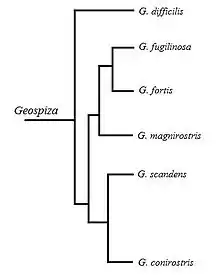
Le genre Geospiza compte 6 espèces, appelées de façon colloquiale les pinsons de Darwin, toutes regroupées sur les îles Galápagos. L'espèce la plus basale de Geospiza est G. difficilis. Les autres espèces sont divisées en deux groupes. Il y a d'abord celles ayant un large bec profond apte à broyer des graines, les « géospizes terrestres » G. fuliginosa, G. fortis et G. magnirostris. Puis, il y a celles ayant un bec affilé plus long qui sert à atteindre les fleurs du cactus et les fruits, les géospizes des cactus G. scandens et G. conirostris[6].
La morphologie très variée du bec au sein de ce groupe est étroitement liée à la structure craniofaciale. La forme spécialisée du bec étant déjà présente à la naissance, cela indique une base génétique durant le développement embryonnaire. L'embryologie de ces espèces démontre qu'à partir du 26e stade embryonnaire, les différences propres à l'espèce quant à la morphologie du bec sont déjà visibles.
Une étude comparative du patron d'expression de différents facteurs de croissance du développement du bec démontre une forte corrélation entre l'expression de BMP4 et la morphologie du bec. Chez les embryons des géospizes ayant un large bec profond, il y a une forte augmentation de l'expression de BMP4 dans le mésenchyme du bec au 26e stade embryonnaire qui se maintient jusqu'au 29e stade. Chez les embryons de G. difficilis et des espèces ayant un long bec affilé, il y a absence de cette augmentation dans l'expression de BMP4.
On en conclut que l'expression accrue et précoce de BMP4 dans le mésenchyme du bec (un exemple d'hétérométrie et d'hétérochronie respectivement) cause une croissance du bec dans l'axe dorso-ventral, ce qui résulte en un bec plus profond et large. La variation de l'expression de BMP4 se limite seulement au bec supérieur. L'expression de cette protéine au bec inférieur demeure constante à travers le genre Geospiza.
Lorsqu'on infecte le mésenchyme du bec d'embryons de poulet avec un vecteur viral dont l'effet est d'augmenter la production locale de BMP4, cela résulte en un bec morphologiquement similaire à celui des « géospizes terrestres ». Inversement, l'infection avec un vecteur viral possédant Noggin résulte en une forte réduction de la forme du bec. On suppose que la régulation de BMP4 dans le mésenchyme du bec découle de l'effet en amont de la protéine sonic hedgehog et du facteur de croissance des fibroblastes 8 (Fgf8), exprimés dans l'épithélium du bec.
Détermination de la morphologie de la mâchoire chez les Cichlidés
Les Cichlidés des Grands Lacs d'Afrique démontrent une diversité surprenante quant à la morphologie de leur mâchoire. Ces lacs regroupent plus de 2 000 espèces issues d'une radiation évolutive. Les espèces qui se nourrissent par broiement possèdent une courte mâchoire avec plusieurs rangées de dents. Cela augmente considérablement l'avantage mécanique de leur mâchoire, soit la transmission de la force lors de l'ouverture et la fermeture de la mâchoire. Chez les espèces qui s'alimentent par succion, la mâchoire possède plutôt une forme élongée avec très peu de rangées de dents, ce qui donne un faible avantage mécanique[8].
Une étude comparative de l'embryologie entre deux Cichlidés du Lac Malawi a été menée pour déterminer l'origine moléculaire de cette diversité morphologique[8]. Les espèces utilisées étaient Labeotropheus fuelleborni, qui possède une courte mâchoire inférieure trapue avec une grande force de broiement, et Metriaclima zebra, qui possède une mâchoire inférieure plus allongée et gracile apte à l'alimentation par succion. L'étude démontre que les différences spécifiques aux espèces quant à la morphologie de la mâchoire ont une forte corrélation avec l'expression de BMP4 au long de l'arc mandibulaire pharyngien. Dans les embryons de M. zebra, l'expression de BMP4 se limitait à la région la plus distale du mésenchyme mandibulaire. Toutefois, l'expression de BMP4 était étendue dans l'ensemble du mésenchyme chez L. fuelleborni. Lorsqu'on provoque une surexpression de BMP4 chez le poisson zèbre Dania rerio, cela cause une croissance de la mâchoire selon l'axe dorso-ventral, donnant une mâchoire plus profonde similaire à celle de L. fuelleborni.
On note que chez les Cichlidés de l'Est de l'Afrique, le taux d'évolution des acides aminés du gène BMP4 est beaucoup plus élevé que celui des autres gènes responsables du développement craniofacial[9]. Ceci témoignerait de la radiation évolutive dont sont issues ces espèces. L'évolution génétique de BMP4 se limite à des substitutions au pro-domaine, ce qui indique des changements dans la régulation de l'expression de BMP4.
Développement de la dentition chez les téléostéens
Les poissons zèbre sont des téléostéens qui ne possèdent pas de dents orales, mais plutôt des dents pharyngiennes. Ce trait, couplé avec l'absence de l'expression de BMP2A, BMP2B et BMP4 dans la région orale au cours du développement embryonnaire, indique une forte corrélation entre les BMP et le développement des dents. L'absence de ces protéines semble être responsable des pertes locales de dents au cours de l'évolution des téléostéens[10].
Régression du pénis chez les oiseaux
Lorsqu'on compare l'organe reproducteur mâle chez deux groupes frères d'oiseaux, Galliformes et Anseriformes (communément appelés les gallinacés et les ansériformes, respectivement), on constate une grande différence entre ces deux groupes[11]. Les gallinacés ne possèdent pas un membre phallique qui s'introduit dans la femelle, mais plutôt un cloaque. Quant aux ansériformes, ils possèdent un phallus bien développé. L'embryologie révèle que les embryons de Galliformes développent un tubercule génital tout comme les Anseriformes à partir du 26e stade embryonnaire. Cependant, vers le 35e stade, il y a arrêt du développement et régression du tubercule par apoptose chez Galliformes. L'apoptose se déclenche au point le plus distal du tubercule.
Lorsqu'on étudie le patron d'expression des facteurs de croissance, on observe chez Galliformes une déviation du patron de BMP4 au niveau du tubercule génital comparativement aux autres groupes. Dans l'embryon du poulet, un gallinacé, il y a surexpression de BMP4 dans le tubercule. Cette surexpression coïncide, à la fois dans le moment et l'emplacement, avec l'apoptose du tubercule, ce qui indique un lien entre les deux phénomènes. L'inhibition de BMP4 par ajout de la protéine Noggin prévient l'apoptose, donnant un organe pénil entièrement développé. Chez les Anseriformes, BMP4 est très peu exprimée au niveau du tubercule. Dans l'embryon du canard, un ansériforme, le fait de provoquer une surexpression de BMP4 dans la région distale du tubercule cause sa régression par apoptose.
Ainsi, la réduction du membre phallique des Galliformes, et même des autres groupes d'oiseaux, ne résulterait pas d'une inhibition de la formation du tubercule génital, mais plutôt d'une apoptose régulée par l'expression de BMP4.
Développement des plumes
L'interaction entre BMP4 et Noggin semble déterminer le processus de développement des plumes chez les oiseaux[12]. Alors que BMP4 promeut la formation du rachis et la fusion des barbes, Noggin promeut le branchement du rachis et des barbes. BMP4 est davantage exprimée dans la région distale du follicule de la plume, tandis que Noggin est exprimé dans les cellules de la pulpe, proche de l'épiderme.
Détermination de l'axe dorsal-ventral
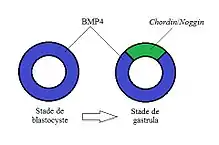
Chez les vertébrés, BMP4 est responsable de l'identité de la région ventrale lors de la gastrulation. Lors du stade de blastocyste, BMP4 est exprimée sur toute la circonférence de l'embryon. Cependant, lors du stade de gastrula, il y a expression de Chordin/Noggin dans la région dorsale, ce qui limite l'expression de BMP4 à la région ventrale[2].
Chez la drosophile, le gène homologue de BMP4 est Dpp (décapentaplégique), responsable de l'identité de la région ventrale au stade de gastrula. Une expérience au cours de laquelle il y a substitution de la région responsable du ligand de Dpp avec celle du ligand de BMP4 humain résulte en un fonctionnement normal du gène Dpp modifié. On obtient tout de même un axe dorsal-ventral normal dans l'embryon de la drosophile, qui résulte en un adulte sain. Cela démontre qu'au cours de l'évolution, il y aurait une forte conservation de la voie de signalisation de BMP4-Dpp[13].
Articles connexes
Références
- (en) Parsons KJ et Albertson RC, « Roles for Bmp4 and CaM1 in Shaping the Jaw: Evo-Devo and Beyond », Annual Review of Genetics, vol. 43, , p. 369-388 (lire en ligne)
- (en) Padgett RW, Wozney JM et Gelbart WM, « Human BMP Sequences Can Confer Normal Dorsal-Ventral Patterning in the Drosophila Embryo », Proceedings of the National Academy of Sciences of the United States of America, vol. 90, no 7, , p. 2905-2909 (lire en ligne)
- « OMIM Entry - * 112262 - BONE MORPHOGENETIC PROTEIN 4; BMP4 », sur www.omim.org (consulté le )
- (en) Urist MR et Strates BS, « Bone morphogenetic protein », Journal of Dental Research, vol. 50, no 6, , p. 1392-1406
- (en) Hogan BLM, « Bone morphogenetic proteins: multifunctional regulators of vertebrate development », Genes & Development, vol. 10, , p. 1580-1594 (lire en ligne)
- (en) Abzhanov A, Protas M, Grant BR, Grant PR et Tabin CJ, « Bmp4 and morphological variation of beaks in Darwin's finches », Science, vol. 305, , p. 1462-1465 (lire en ligne)
- (en) Abzhanov A, Kuo WP, Hartmann C, Grant BR, Grant PR et Tabin CJ, « The calmodulin pathway and evolution of elongated beak morphology in Darwin's finches », Nature, vol. 442, , p. 563-567 (lire en ligne)
- (en) Albertson RC, Streelman JT, Kocher TD et Yelick PC, « Integration and Evolution of the Cichlid Mandible: The Molecular Basis of Alternate Feeding Strategies », Proceedings of the National Academy of Sciences of the United States of America, vol. 102, no 45, , p. 16287-16292 (lire en ligne)
- (en) Terai Y, Morikawa N et Okada N, « The Evolution of the Pro-Domain of Bone Morphogenetic Protein 4 (Bmp4) in an Explosively Speciated Lineage of East African Cichlid Fishes », Molecular Biology and Evolution, vol. 19, , p. 1628-1632 (lire en ligne)
- (en) Wise SB et Stock DW, « Conservation and divergence of Bmp2a, Bmp2b, and Bmp4 expression patterns within and between dentitions of teleost fishes », Evolution & Development, vol. 8, no 6, , p. 511-523 (lire en ligne)
- (en) Herrera AM, Shuster SG, Perriton CL et Cohn MJ, « Developmental Basis of Phallus Reduction during Bird Evolution », Current Biology, vol. 23, , p. 1065-1074 (lire en ligne)
- (en) Yu M, Wu P, Widelitz RB et Chuong CM, « The morphogenesis of feathers », Nature, vol. 420, , p. 308-312 (lire en ligne)
- (en) Holley SA et Ferguson EL, « Fish are like flies are like frogs: conservation of dorsal-ventral patterning mechanisms », BioEssays, vol. 19, no 4, , p. 281-284 (lire en ligne)
 Portail de la biologie
Portail de la biologie