Cohésine
La cohésine est un complexe qui maintient les deux chromatides sœurs associées après la duplication jusqu'à l'anaphase
Structure
Le complexe cohésine est conservé de la levure à l’homme. Il consiste en au moins cinq sous-unités formant un grand anneau protéique, d’un diamètre interne de 35 nm (pour comparaison, le diamètre d’une fibre de chromatine est de 10 nm).
Association de la cohésine avec les chromosomes
L’hypothèse communément admise est que cet anneau encerclerait topologiquement les deux chromatides sœurs. En effet, l’ouverture de l’anneau par clivage (expérience in vitro et in vivo) détruit l’association avec la chromatine, et réciproquement, le clivage de l’ADN libère les cohésines de la chromatine (ce qui est en faveur d'une interaction topologique plutôt que physique).
Ancrage
La cohésion entre les deux chromatides sœurs opère dès la phase S de réplication via la cohésine. Cela suppose l’ancrage de la cohésine avant cette étape. La cohésine est globalement présente sur toute la longueur du chromosome mais sa répartition demeure inégale. En particulier, le centromère et sa région alentour est la plus riche en cohésine. Cette particularité est plus qu’anecdotique puisqu’il a été établi que la cohésion péricentromèrique contribue à l’orientation et la ségrégation normale durant la méiose.
Encerclement
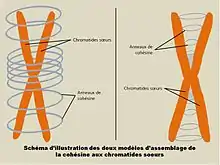
En considérant que la cohésine maintient les deux chormatides sœurs en les encerclant, plusieurs interprétations peuvent être avancées concernant la manière dont les chromatides se sont retrouvées toutes deux encerclées. Un premier modèle envisage qu’après s’être ancré sur la chromatine, l’anneau de cohésine encercle un premier brin d’ADN. À la suite de la réplication d’ADN pendant la phase S du cycle cellulaire, le deuxième brin se retrouve de fait encerclé dans l’anneau. Le deuxième modèle consiste à envisager les facteurs de réplication comme facteurs facilitant l’ouverture de l’anneau.
Libération des chromosomes
L’élimination totale de cohésine le long des brins d’ADN est la résultante du clivage du complexe protéique au niveau d’une de ses sous-unité par l’enzyme séparase. Dans la mitose cette sous-unité est appelée Scc1 tandis que dans la méiose il s’agit d’une sous-unité homologue appelée Rec8.
Fonction
La cohésion des chromatides sœurs est cruciale dans le bon déroulement de la mitose et de la méiose.
1. Mitose
La cohésine maintient les deux chromatides sœurs ensemble jusqu'à l'entrée de l'anaphase, où à la suite de la dégradation de celle-ci, les deux chromatides se séparent et migrent vers les pôles opposés de la cellule. La solidarité entre les deux chromatides sœurs joue un rôle important lors de la métaphase en ce qu'elle promeut l'attachement de chaque kinetochore à des pôles opposés. Cette « bi-orientation » permet une division normale: au contraire, si les deux chromatides migraient dans la même direction (« mono-orientation »), la division résulterait en des cellules filles aneuploïdes.
En métaphase, chaque chromosome est attaché de façon bilatérale via ses kinétochores aux microtubules du fuseau. La cohésine oppose une résistance à la traction exercée par les microtubules se liant aux kinétochores de chaque chromatide. La cohésion des chromatides sœurs compense la force de traction exercée par les microtubules. La tension générée via la cohésion des deux chromatides sœurs est cruciale dans le bon déroulement de l'anaphase pour deux raisons :
- La tension permet la stabilisation de la liaison microtubule-kinétochore. Elle favorise la bi-orientation des deux chromatides sœurs, c'est-à-dire leur migration vers des pôles opposés ;
- La tension générée aux kinétochores indique au checkpoint mitotique que chaque chromatide est attachée de façon correcte au fuseau et que l’anaphase peut avoir lieu. Ce mécanisme permet de rendre le déclenchement de l’anaphase dépendant de l’attachement bilatéral correct de tous les chromosomes. Plusieurs études se sont penchées sur la manière dont la tension influençait la mise en route ou non des systèmes de correction d'erreurs. Un des mécanismes proposés met en jeu la kinase Aurora B, enzyme clée dans le checkpoint mitotique. En l'absence de tension, une sous unité de la kinase, appelée Ipl1, inhiberait les interactions entre kinétochores et centromères via la phosphorylation de nombreux substrats situés sur la face externe des kinétochores. Ainsi la machine de correction devrait intervenir pour permettre un attachement correct entre kinétochores et microtubules. À l'inverse, lorsqu'une tension est générée, Ipl1 située au niveau de la face interne des kinétochores, se verrait physiquement séparée de ses substrats situés sur la face externe, annulant ainsi la phosphorylation ainsi que l'intervention de la machine de correction d'erreur.
Ce mécanisme permet de rendre le déclenchement de l’anaphase dépendant de l’attachement bilatéral correct de tous les chromosomes. Par ailleurs, la cohésine joue un rôle crucial dans la mitose en ce qu'elle assure la réparation par recombinaison homologue de cassures double brin d’ADN, car la chromatide sœur intacte sert de matrice à la réparation.
2. Méiose
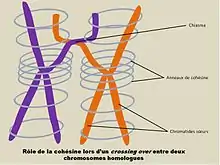
La méiose comprend non seulement la séparation des chromatides sœurs lors de l’anaphase II mais aussi la séparation, lors de l’anaphase I, des chromosomes homologues étroitement appariés dès la prophase I. La cohésine joue un rôle à ces deux niveaux. La cohésine contribue à la cohésion des chromosomes homologues jusqu’à anaphase de la méiose I. Lorsqu’un enjambement (crossing over) a lieu entre deux paires de chromosomes homologues dès la prophase I, l’entremêlement réalisé implique que la cohésion le long des bras des chromatides résulte en une cohésion des deux chromosomes homologues. Il ne s'agit pas là de la cohésion au niveau du chiasma mais de l'action de la cohésine, le long des chromatides sœurs, alors que des chromatides non sœurs s'entremêlent. A l'anaphase de la méiose II, la cohésine joue un rôle analogue à celui joué dans la mitose puisqu’elle permet la cohésion des chromatides sœurs jusqu’à l’anaphase II, puis concourt à leur orientation vers des pôles bien distincts.
De la séparation des chromosomes homologues à la séparation des chromatides sœurs
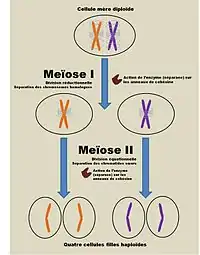
La séparation des chromatides sœurs résulte du clivage de la cohésine par l’enzyme séparase. Cette séparation a lieu après une première séparation, celle des chromosomes homologues, qui a également nécessité le clivage de la cohésine. La particularité de la cohésine dans la méiose est qu’elle n’est pas totalement éliminée du premier coup: elle persiste au niveau du centromère et de la région périphérique du centromère (péri-centromère) jusqu’à l’anaphase II avant d’être totalement éliminée des chromosomes.
Cette particularité permet à la méiose d’être distinguée de façon binaire (méiose I et méiose II) et de former 4 cellules filles haploïdes à la suite d’une première division réductionnelle, puis d’une division équationnelle. L’élimination en 2 étapes de la cohésine sur toute la longueur des chromatides sœurs s’explique par la présence de la sous-unité Rec8, spécifique de la méiose. Celle-ci, tout comme son homologue Scc1 de la mitose correspond à la partie où s’opère le clivage via l’enzyme séparase, cependant elle diffère par sa capacité à résister à la séparase au niveau du péri-centromère. Plusieurs points de vue peuvent être adoptés pour expliquer quelle propriété spécifique de Rec8 permet sa conservation au niveau du péri-centromère. Notamment, le fait que Rec8 ait besoin d’être phosphorylé pendant la méiose 1 peut être mis en relation avec l’action de la protéine Shugoshin (Sgo1) située dans la région du péri-centromère. Dans cette optique, en favorisant la formation d’un certain complexe protéique clé pour éviter la phosphorylation de Rec8, cette protéine joue un rôle déterminant dans la résistance de l’anneau cohésine autour des régions proches du centromère. D’autres agents contribuent à réguler la protection de la cohésine au niveau du péri-centromère, notamment la protéine Spo13, spécifique de la méiose ou encore la kinase Aurora Ipl1.
Migration des chromosomes homologues
De façon analogue à la migration des chromatides sœurs telles que décrites pour la mitose, le checkpoint mitotique liée à la génération de tension via la cohésine joue un rôle clé dans l’orientation diamétralement opposée des chromosomes homologues. Cependant, quels sont les moyens que la cellule a pour sentir la tension et réagir en conséquence est un questionnement ouvert. En effet, les procédés mis en jeu ne sont pas tout à fait assimilables à ceux mis en jeu lors de l’orientation des chromatides sœurs.
En médecine
Les mutations sur l'un des gènes codant l'une des protéines des cohésines entraînent des « cohésinopathies ».
Voir aussi
Bibliographie
Marston Adèle L. Chromosome Segregation in Budding Yeast: Sister Chromatid cohesion and Related Mecanisms. Genetics Society of America,
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire  Portail de la médecine
Portail de la médecine