Cycle de l'azote
Le cycle de l'azote est un cycle biogéochimique qui décrit la succession des modifications subies par les différentes formes de l'azote neutre en formes réactives (diazote, nitrate, nitrite, ammoniac, azote organique) et vice-versa.

Les analyses isotopiques de l'azote faites dans différents compartiments géologiques et de la biosphère et notamment dans les sédiments lacustres montrent que ce cycle a été récemment fortement perturbé par l'Homme qui a plus que doublé la quantité d'azote réactif (Nr) annuellement ajoutés à la biosphère[1], essentiellement à partir de 1895 ± 10 ans (± 1 pour l'écart-type) avec une forte augmentation dans les années 1960 à 2010, principalement dans l'hémisphère Nord[2].
Généralités
L'azote est un composé de base de la matière organique, en particulier pour la synthèse des acides aminés constituant les protéines et les bases nucléiques de l'ADN. De ce fait, la réduction de N2 est considérée comme aussi essentielle à l'évolution et à la perpétuation de la vie sur Terre que la photosynthèse[3].
D'autre part, les composés de l'azote participent au métabolisme cellulaire, comme matériau à oxyder en milieu oxygéné, ou au contraire comme oxydant accepteur d'électrons en milieu anoxyque.
Cycle biologique
Dans le cycle biologique, l'azote se présente sous forme d'ion ammonium, de nitrate ou de nitrite, ou de matière organique.
- Les végétaux assimilent l'azote, soit directement sous forme d'ion ammonium, soit sous forme de nitrate ou de nitrite qu'ils réduisent par photosynthèse ; et le transforment en matière organique.
- Les champignons, animaux et monocellulaires hétérotrophes ne peuvent assimiler l'azote que sous forme de matière organique, qu'ils utilisent également comme source d'énergie.
- La décomposition de matières organiques restitue de l'ammoniaque.
- L'oxydation de cet ammoniaque par certaines bactéries leur fournit de l'énergie, et produit des nitrites, puis des nitrates.
Cependant, dans notre environnement globalement oxydant, l'ion ammonium est normalement oxydé et ne s'accumule pas de manière significative. En dehors de la biomasse, où son nombre d'oxydation est -3, l'azote du cycle biologique se présente généralement sous forme oxydée, N(+iii) ou N(+v)
Cycle planétaire
Ce cycle biologique s'insère dans un cycle plus global :
- Quelques bactéries et cyanobactéries peuvent fixer l'azote atmosphérique sous forme de NH3 incorporé, et sont le point d'entrée du cycle biologique ;
- En milieu anaérobie, d'autres bactéries réalisent la dénitrification du milieu en les réduisant successivement en monoxyde d'azote, puis en protoxyde d'azote, et enfin en diazote, qui est restitué à l'atmosphère et sort du cycle biologique.
Le cycle naturel de l'azote a toutefois été fortement perturbé par les activités humaines, qui ont modifié la disponibilité de l'azote, avec des implications encore mal évaluées pour les écosystèmes à travers le monde[réf. souhaitée]. De nos jours, près de 30% de l'azote fixé est produit industriellement par le procédé Haber[4].
Imbrication des cycles biologiques
À travers la fixation biologique de l'azote réalisée par certaines cyanobactéries, l’inhibition de la nitrogénase par l'O2, que ce soit directement ou indirectement via des radicaux oxydants, constitue un régulateur fort de la teneur en oxygène dans l'atmosphère, dans le cycle de l'oxygène, par la rétroaction négative qu'il a sur la fixation de l'azote[5]. Une inhibition dans la fixation de l'azote conduirait à une chute de la productivité primaire, à la fois directement pour les cyanobactéries diazotrophes et indirectement pour l'ensemble de la productivité primaire de l'hydrosphère, induisant à plus long terme, pour le cycle du carbone, une réduction dans le rythme d'enfouissement du carbone organique. Sans enfouissement il n'y a pas de création nette d'O2 par la biosphère, et l'oxygène en excès espèce très réactive, disparaît progressivement par l'oxydation de la surface de la planète.
Étapes du cycle biologique
Fixation

Le cycle part de l'azote atmosphérique. L'atmosphère contient une grande quantité de diazote (79 % en volume), mais cet azote gazeux est très peu biodisponible pour la plupart des espèces.
Seules quelques bactéries et cyanobactéries (principalement présentes dans le sol), telles qu'Azotobacter vinelandii, peuvent fixer l'azote atmosphérique, sous forme de NH3 incorporé, qu'elles transforment ensuite en matière organique. Certaines de ces bactéries produisant de l'ammoniaque, comme Rhizobium, vivent en symbiose avec des plantes.
Les bactéries fixatrices de l'azote produisent de l'ammoniaque (NH4OH) à partir de l'azote atmosphérique et de l'hydrogène de l'eau (l'ammoniaque est le nom de la forme soluble dans l'eau du gaz ammoniac). La fixation de l'azote nécessite un flux important d'ATP[3]. Le processus de fixation d'une molécule d'azote en deux molécules d'ammoniac est couplé à l'hydrolyse de 16 ATP en milieu acide, et cogénère une molécule d'hydrogène. La réaction de fixation est globalement la suivante[6] :
Elles le font grâce à une enzyme, la nitrogénase qui est capable de casser la triple liaison N≡N qui rend l'azote gazeux si stable, mais uniquement dans certains contextes, car cette réaction de transformation d'azote atmosphérique en NH3 assimilable par les organismes a un coût énergétique et métabolique élevés, et fait appel à d'autres éléments nécessaires à la biosynthèse de la nitrogénase. Pour ces raisons, l'azote biodisponible est un puissant facteur limitant pour la production de biomasse algale et végétale, et donc pour la productivité de nombreux écosystèmes. Ceci explique aussi que la disponibilité en azote joue un rôle majeur dans la structuration et les dynamiques d'évolution de nombreuses communautés végétales.
La décomposition d'organismes morts (nécromasse) par des bactéries saprophytes sous forme d'ions ammonium (NH4+) est une autre source d'ammoniaque.
Nitritation et nitratation
.jpg.webp)
Dans la nature, la décomposition de matières organiques restitue de l'ammoniac, qui donne par exemple une odeur d'égout aux eaux trop stagnantes.
Dans les sols bien oxygénés, mais aussi en milieu aquatique oxygéné, des bactéries transforment l'ammoniac (NH3) en nitrite (NO2−), puis en nitrates (NO3−), au cours du processus de nitrification. L'oxydation de cet ammoniac par ces bactéries leur fournit l'énergie nécessaire pour transformer le CO2 en matière organique (autotrophie)[7].
La nitrification se déroule en deux étapes[8], réalisées par deux populations bactériennes différentes. Une première étape de nitritation oxyde l'amoniac en nitrite, consommant une molécule d'oxygène par molécule formée :
Le déchet toxique des nitrites de la première communauté bactérienne est ensuite utilisé comme métabolite par la seconde. Ce phénomène de symbiose est appelé syntrophie[9]. Cette seconde étape de nitratation transforme ensuite le nitrite en nitrate:
La nitrification est une source d'acidité pour le sol. C'est cette consommation en milieu oxygéné qui est à l'origine de la présence de nitrates assimilables par les végétaux.
En aquariophilie, la nitrification est indispensable pour que l'aquarium ne s'empoisonne pas sous l'effet de l'accumulation de l'ammoniaque. Pour favoriser la nitrification, on fait circuler l'eau sur un « filtre » dont la surface fournit un support aux bactéries. À la fin du cycle, restent les nitrates qui servent de nourriture aux plantes, et qui en excès peuvent favoriser la prolifération des algues, voire s’avérer problématiques pour les poissons eux-mêmes.
Nitratation alternative par l'ozone

Les éclairs dissocient les molécules d'oxygène et d'azote dans l'atmosphère, formant des atomes exités qui peuvent se recombiner sous forme de NO et NO2, et d'ozone O3. Ces trois espèces sont en équilibre mutuel, le NO2 étant photo-dissocié en NO en produisant de l'oxygène atomique, lequel se combine à de l'oxygène pour former de l'ozone (en présence d'une molécule M stabilisante), lequel forme à nouveau du NO2 :
- NO2 + hν → NO + O*
- O* + O2 (+ M) → O3 (+ M)
- O3 + NO → NO2 + O2
Le NO2 peut également réagir avec l'ozone pour former du NO3, qui est rapidement dissocié par photolyse pendant le jour :
- NO2 + O3 → NO3 + O2 et NO3 + hν → NO2 + O*
De nuit, cependant, il peut se combiner à une autre molécule de NO2 pour former du pentoxyde d'azote :
- NO2 + NO3 (+ M) → N2O5 (+ M)
Le pentoxyde d'azote se dissout dans l'eau en phase liquide (dans les nuages ou les aérosols) où il réagit pour former de l'acide nitrique :
- N2O5 + H2O → 2 HNO3
Cet acide retombe au sol sous forme de pluie acide, et contribue à la fertilité du sol sous forme de nitrate NO3−
Assimilation


Les végétaux peuvent assimiler l'azote, soit sous forme d'ion ammonium, soit sous forme d'ion nitrate (NO3−) et de nitrite (NO2−) d'autre part. Ces deux dernières formes d'azote inorganiques sont d'abord converties en ammonium, puis en glutamine, puis ce dernier est transformé en matière organique. La réduction préalable des nitrates nécessite de l'énergie qui provient essentiellement de la photosynthèse.
Les organismes comme les plantes, les champignons et certaines bactéries qui ne peuvent pas fixer l'azote (N2) dépendent de leur capacité à assimiler le nitrate ou l'ammoniac pour leurs besoins.
Les végétaux pour se nourrir absorbent (au travers des feuilles pour certaines plantes aquatiques et sinon via les racines) les ions nitrate (NO3−) et, dans une moindre mesure, l'ammonium NH4+ présent dans le sol.
- Réduction des nitrates
Dans un premier temps, le nitrate est converti en nitrite par la nitrate réductase :
Le nitrite est ensuite réduit en ammoniac par la ferrédoxine-nitrite réductase :
- NO2− + 6 ferrédoxine réduite + 7 H+ NH3 + 2 H2O + 6 ferrédoxine oxydée

L'assimilation par transformation d'un nitrate en ammoniac consomme un ion H+, qui doit être équilibré par l'expulsion d'un OH− qui tend à rendre le sol alcalin.
- Assimilation de l'ammoniac
L'assimilation proprement dite porte sur l'ammoniac, qu'il soit absorbé directement ou créé par réduction.
Dans les chloroplastes, la glutamine synthétase incorpore cet ammoniac dans le cadre de la photorespiration, transformant un glutamate en glutamine :
- NH3 + L-glutamate + ATP L-glutamine + ADP + Pi.
Le glutamate joue un rôle essentiel dans le cycle de l'azote chez les animaux et les microbes, servant à la fois d'accepteur (comme ici) et de donneur pour les transferts d'ammoniac dans le cycle.
Formation et destruction des protéines
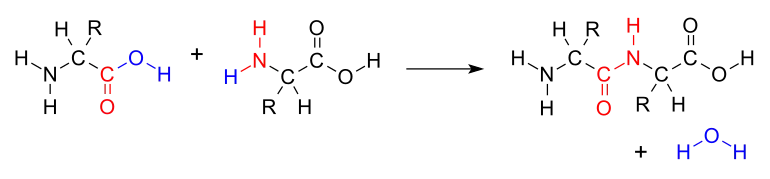
L'assimilation de l'azote, à partir de composés azotés inorganiques présents dans l'environnement, permet ensuite la formation de composés azotés organiques tels que les différents acides aminés, par transfert du groupe amine d'une chaîne à l'autre, puis les protéines qui en sont formées par polymérisation via la liaison peptidique.
Les organismes qui ne peuvent pas assimiler le nitrate ou l'ammoniac pour leurs besoins, comme les animaux, dépendent entièrement de l'azote organique provenant de leur nourriture. Les végétaux constituent ainsi la source primaire d'azote assimilable par les animaux.
Les protéines digérées sont inversement décomposées en leurs acides animés par protéolyse (ou dégradation protéique), qui détruit les liaisons peptidiques par hydrolyse. Les acides aminés ainsi libérés peuvent être réutilisés dans l'organisme pour de nouvelles synthèses protéiques, et rester dans la biosphère, ou être irréversiblement dégradés par oxydation pour être utilisées comme source d'énergie.
Catabolisme des acides aminés
La dégradation des acides aminés est réalisée chez les animaux ou champignons ou bactéries qui se nourrissent de matière organique. Cette digestion convertit l'azote de la matière organique en ammoniac.
Elle commence par une « désamination », séparant le groupement alpha aminé du squelette carboné pour le transférer sur l'acide glutamique. La réaction est catalysée par une transaminase.
Le L-glutamate ainsi produit subit ensuite une désamination oxydative, catalysée par la glutamate déshydrogénase, qui élimine finalement le groupement α-aminé sous forme de NH4+. Chez les mammifères, l'ion ammonium, toxique pour la cellule, rentre ensuite dans le cycle de l'urée pour être excrété. Suivant les animaux, l'excrétion peut prendre la forme alternative d'ammoniac, d'acide urique ou de créatinine.
De son côté, le squelette carboné est ensuite oxydé, ce qui correspond à la fonction respiratoire du cycle de l'oxygène.
Dénitrification
En milieu anaérobie, d'autres bactéries utilisent les nitrates et nitrites pour la respiration cellulaire comme accepteur d'électron[10], et réalisent la dénitrification du milieu en les réduisant successivement en monoxyde d'azote, puis en protoxyde d'azote, et enfin en diazote, qui est restitué à l'atmosphère et sort du cycle biologique.
La dénitrification réduit l'azote en un état gazeux N2 ou N2O, inassimilable par les plantes, et le fait sortir du cycle biologique.
En milieu anoxique (sol ou milieu aquatique non oxygéné), des bactéries dites dénitrifiantes transforment les nitrates en gaz diazote, c'est la dénitrification. La dénitrification suit tout ou partie de ces réactions successives (l'enzyme catalysant la réaction figurant entre parenthèses) :
- NO3− + 2 H+ + 2 e−→ NO2− + H2O (nitrate réductase)
- NO2− + 2 H+ + e− → NO + H2O (nitrite réductase)
- 2 NO + 2 H+ + 2 e− → N2O + H2O (oxyde nitrique-réductase)
- N2O + 2 H+ + 2 e− → N2 + H2O (oxyde nitreux-réductase)
Le processus complet peut s'exprimer sous forme d'une réaction redox équilibrée, par laquelle le nitrate (NO3−) est entièrement réduit sous forme d'azote N2 :
- 2 NO3− + 10 e− + 12 H+ → N2 + 6 H2O
Les bactéries qui effectuent le processus d'anammox appartiennent au phylum bactérien des planctomycètes, dont les genres les mieux connus sont Planctomyces et Pirellula. Dans ce processus biologique, le cation ammonium NH4+ est oxydé par l'anion nitrite NO2− pour donner du diazote N2. Ce processus contribue à hauteur de 50 % de l'azote produit dans les océans. C'est donc un important puits d'azote qui limite ainsi la production primaire des océans. La réaction globale s'écrit :
Évolution du cycle
D'une manière ou d'une autre, l'assimilation de l'azote pour former de la biomasse ne se fait que sous forme NO3-, et la biosphère dépend de la formation de cet ammoniac, une des formes les plus réduites de l'azote. Par ailleurs, l'azote sous différents degré d'oxydation fait l'objet de transformations variées dans la biosphère, généralement pour en tirer de l'énergie.
Le cycle de l'azote tel qu'on le connaît aujourd'hui est largement associé à la biosphère, et dépend de l'oxygène pour l'étape cruciale qu'est la nitrification biologique (ou celle alternative par l'ozone). Cependant, cette étape n'a pas pu exister dans la chimie pré-biotique, qui se place dans un contexte sans oxygène et largement réducteur. Dans l'histoire récente de la planète, la première étape biologique du cycle de l'azote est la fixation de l'azote dans une réaction complexe faisant intervenir la nitrogénase ; mais l'origine évolutive de cette enzyme est mal connue et fait débat[11].
Antérieurement à la Grande Oxydation, le cycle de l'azote a dû être très différent de ce que l'on constate aujourd'hui[11]. Sans oxygène, le « cycle de l'azote » n'est pas un cycle, mais une voie à sens unique vers les formes réduites[11]. Le cycle actuel est nécessairement récent, et postérieur à la Grande Oxydation.
À l'origine de la vie, il n'y avait ni oxygène libre, ni nitrogénase. L'azote devait nécessairement être directement présent sous forme NO3-. En l'absence de métabolisme, il n'a pu être rendu disponible qu'à travers des transformations abiotiques permettant de convertir l'azote disponible, majoritairement sous forme de N2 dans l'atmosphère. Il s'agissait probablement par des réactions transformant le N2 en oxyde d'azote et acide nitrique sous l'effet de la foudre ou de la photolyse, dans une atmosphère par ailleurs chargée de CO2 et de H2O ; mais ces réactions chimiques sont mal comprises et sujettes à débat[11]. De plus, si l'océan primitif était riche en fer ferreux, celui-ci réagit rapidement avec les oxydes d'azote, pour s'oxyder en fer ferrique et les réduire en ammoniaque[11].
En tout état de cause, d'une manière ou d'une autre, l'origine de la vie s'est faite en présence de NO3- immédiatement disponible, et les premières cellules ont pu se reproduire.
Cette forme de vie a rencontré une crise avec l'invention de la photosynthèse oxygénique, qui a fait progressivement disparaître le NO3- présent dans les océans. Cette pénurie sur un composant essentiel de la vie primitive a réduit la biosphère (et par voie de conséquence régulé la production d'oxygène), le NO3- étant devenu un facteur limitant dans la compétition aux ressources. De ce fait, la capacité à catalyser la réduction du N2 en NH3 a constitué un avantage adaptatif, poussant vers l'émergence d'une nitrogénase de plus en plus performante.
Avec ce nouvel accès à l'azote, la biosphère a pu reprendre son expansion, notamment pour les bactéries responsables de la photosynthèse oxygénique, qui n'ont plus été limitées par l'absence d'azote fixé. Avec l'apparition de la nouvelle atmosphère d'oxygène, le cycle moderne a pu se mettre en place.
Azote géologique
Une partie souvent oubliée du cycle de l'azote passe par le substratum rocheux là où de l'azote était présent dans la matière organique sédimentée ou liée à cette dernière ; de l'« azote géologique » est aussi trouvé dans certaines eaux thermales, provenant alors de roches sédimentaires, du manteau et parfois de sources météoriques[12].
Les taux d'azote dans les roches varient de simples traces (<200 mg N/kg) dans les granits à des taux écologiquement importants, dépassant 1 000 mg N/kg dans certaines roches sédimentaires et métasédimentaires[13]. Des roches riches en azote sont présentes un peu partout dans le monde et pourraient aussi jouer un rôle local, sous-estimé, dans la disponibilité en azote pour le réseau trophique. En fait, selon Schlesinger (1997) près de 20 % de la totalité du cycle de l'azote serait fixé dans les roches[14].
Ce rôle pourrait avoir été négligé en raison d'un manque de méthodes analytiques de routine pour la quantification de l'azote dans les roches. Les dépôts de nitrates accumulés dans les régions arides et semi-arides sont notamment des réservoirs potentiellement importants[12].
Dans la plupart des cas, les taux élevés d'azote dans l'eau et le sol ont une origine humaine. Parfois, ils peuvent être attribués aux effets d'intempéries sur le substrat rocheux (en cas de roche-mère riche en ammonium), avec dans certains environnements, une quantité d'azote libérée du socle rocheux supérieure aux besoins des plantes, et pouvant contribuer à saturer le milieu terrestre et l'écosystème local en produisant des phénomènes d'eutrophisation et de dystrophisation. L'oxydation du nitrate d'ammonium peut aussi contribuer à l'acidification des sols et au phénomène plus général d'acidification des eaux douces, susceptible d'inhiber la revégétalisation et renaturation spontanée de certains écosystèmes[12].
Il semble que les écosystèmes établis sur des roches à teneurs élevées en azote soient plus vulnérables à l'anthropisation ; plusieurs études récentes ont montré que l'azote géologique existe en quantité significative, qu'il peut être libéré et réactivé dans certaines circonstances, avec alors selon Holloway et Dahlgren (2002) « un potentiel d'amplification des impacts humains sur le cycle de l'azote dans les écosystèmes terrestres et aquatiques »[12].
L'azote géologique et inorganique (outre les formes organiques de l'azote ou liée à la matière organique, surtout trouvées dans les sédiments organiques jeunes) est notamment constitué des minéraux suivants :
| Minéral | Formule | Occurrence (exemples) |
|---|---|---|
| Osbornite | TiN | Chondrite[15] |
| Sinoite (en) | Si2N2O | Chondrites[16],[17] |
| Nitratine | NaNO3 | Désert d'Atacama (Chili) ; désert de Mojave (États-Unis) ; Ouest des États-Unis ; grottes (États-Unis)[18],[19],[20] |
| Nitre | KNO3 | Désert d'Atacama (Chili), Ouest des États-Unis ; grottes (États-Unis)[18],[20]... |
| Nitrocalcite | Ca(NO3)2·4H2O | Dépôts dans des grottes, AZ, États-Unis[21] |
| Svéite | KAl7(NO3)4Cl2(OH)16·8H2O | Dépôts dans des grottes (ex : Venezuela)[22] |
| Gerhardtite | Cu2(NO3)(OH)3 | Dépôts liés au minerai de cuivre (ex : Australie ; Congo ; AZ, États-Unis)[23],[24],[25] |
| Likasite | Cu3(OH)5(NO3)·2H2O | Dépôts liés au minerai de cuivre (Congo)[26] |
| Darapskite | Na3(SO4)(NO3)·H2O | Origine hydrothermale (Antarctique)[27] ; Caliche : Désert d'Atacama (Chili)[18] ; désert de Mojave (États-Unis)[18] |
| Humberstonite | Na7K3Mg2(SO4)6(NO3)2·6H2O | Caliche : Désert d'Atacama (Chili), Désert de Mojave (États-Unis)[28],[19] |
| Ammoniojarosite | (NH4)Fe3+(SO4)2(OH)6 | Minerais : WY & UT, États-Unis ; origines hydrothermales Geysers, CA[29],[30],[31] |
| Boussingaultite | (NH4)2Mg(SO4)2·6H2O | Origines hydrothermales Geysers (CA)[32],[33] |
| Lecontite | (NH4,K)Na(SO4)·2H2O | Évaporites (ex : Pakistan)[34] |
| Letovicite | (NH4)3H(SO4)2 | Origine hydrothermale : Geysers CA, États-Unis ; Campi Flegrei (Italie)[31],[35] |
| Mascagnite | (NH4)2SO4 | Origine hydrothermale : Geysers, CA, États-Unis[31] |
| Salmiac | (NH4)Cl | Source hydrothermale (ex : Etna et Vésuve en Italie)[36] |
| Tschermigite | (NH4)Al(SO4)2·12H2O | Source hydrothermale ; Geysers, CA. États-Unis ; Campi Flegrei en Italie ; Taupo en Nouvelle-Zélande[31],[35],[37] |
| Mundrabillaite | (NH4)2Ca(HPO4)2·H2O | Dépôts dans des grottes (Australie)[38] |
| Ammonian fluorapophyllite | (NH4,K)Ca4Si8O20(F,OH)·8H2O | Source hydrothermale : Calvinia (Afrique du Sud) et Guanajuato (Mexique)[39] |
| Ammonioleucite | (NH4)AlSi2O6 | Roches métamorphiques volcaniques : Japon[40] ; Source hydrothermale : Geysers, CA[41] |
| Buddingtonite | NH4AlSi3O8 | Source hydrothermale : Sulphur Bank Mine, CA ; Cedar Mt, NV, États-Unis ; Japon ; Phosphoria Fm.: ID, États-Unis ; oil shale : Australia[42] ,[43],[44],[45],[46] |
| Tobélite | (NH4,K)Al2(Si3Al)O10(OH)2 | Argiles hydrothermiquement altérées : Japon; UT, États-Unis ; pétrole de schiste en mer du nord[47],[48],[49] |
En mer
Sous forme de nitrate ou d'ammoniaque, l'azote est très soluble dans l'eau et ainsi très mobile dans les écosystèmes. Le ruissellement, l'érosion et les pluies, trois phénomènes exacerbés par l'artificialisation des milieux et/ou modifiés par le réchauffement climatique tendent à ramener les nitrates non captés par la biomasse terrestre vers les lacs et surtout vers les nappes phréatiques et les océans. Dans la mer, comme sur terre, l'azote dissous dans l'eau est capté (via la photosynthèse) par les plantes et certaines bactéries, puis concentré dans le réseau trophique sous forme de protéines animales notamment. Le cycle de l'azote est particulièrement perturbé dans les zones marines littorales et estuariennes[50].
Tout poisson et tout animal marin concentre dans sa biomasse (matière organique) une partie de l'azote accumulé par les végétaux ou animaux qu’il a consommés, lesquels l’ont prélevé dans le stock océanique, alimenté par l’atmosphère, et de plus en plus excessivement par les apports terrigènes (engrais, et eaux usées essentiellement).
Depuis le développement de la pêche industrielle et intensive, cette quantité n’est plus négligeable. En effet, chaque poisson pêché en mer correspond donc à un retrait d’azote du compartiment océanique. Cette quantité a été évaluée pour 58 écosystèmes marins répartis sur le globe par une étude[51] récente sur le cycle de l'azote dans les régions côtières. En 1960, la pêche en mer ramenait vers la terre 60 % des composés azotés estimés être apportés par les fertilisants lessivés par les pluies. Or, de 1960 à 2004, ces apports azotés ont été multipliés par 7,5, alors qu’en raison d’une raréfaction des espèces commercialisables à la suite de la surexploitation des océans, les prises de pêche n’ont augmenté que de 2,5. Seulement environ 20 % de l’azote perdu en mer par l’agriculture sont donc en 2008 « récupérés » par les activités de pêche, lesquelles devraient être pour ces raisons mieux prises en compte dans les calculs des bilans d’azote et risques d’eutrophisation.
Une interprétation facile et rapide de ces résultats serait de croire qu’en augmentant encore la pêche on diminuerait l’eutrophisation marine. Cette piste serait illusoire, rappelle Greenpeace l'auteur de l'étude elle-même, notamment parce que poursuivre la surpêche conduira rapidement à la disparition pure et simple de la plupart des espèces commerciales, et donc à une eutrophisation pire encore. La solution préconisée par les ONG environnementales et de nombreux organismes scientifiques, et notamment par un rapport récent[52] du Conseil de la recherche portant sur la « qualité de l'eau du fleuve Mississippi au regard de la Loi sur l'eau : progrès, défis et opportunités ». Ce rapport recommande que l'EPA et l'USDA coopérèrent mieux pour réduire les impacts de l'agriculture sur la qualité de l'eau du Mississippi et du nord du golfe du Mexique (zone morte de 22 000 km2 environ en 2007-2008 ; la plus vaste du monde). L'excès d'azote cause la prolifération de phytoplancton qui se décompose en consommant l'oxygène dissous dans l'eau aux dépens des organismes vivants qui disparaissent alors[53]. La seule solution durable serait de limiter la quantité de pertes d’engrais azotés d'origine agricole et de reconstituer un stock de poisson suffisant pour alimenter une pêche durable. Les indicateurs disponibles laissent penser que ces deux options peinent à se mettre en place.
Dans un aquarium

1 - Nourriture et nutriments
2 - Production d'urée et d'ammoniac
3 - Ammoniac → nitrites (Nitrosomonas)
4 - Nitrites → nitrates (Nitrospira)
5 - Évaporation
6 - Lumière
7 - Sol
8 - Oxygène
9 - Dioxyde de carbone
Dans un aquarium, ce sont les déjections des poissons et la nourriture non consommée qui sont à l'origine de la formation d'ammoniac. Ce produit extrêmement toxique pour les animaux aquatiques est transformé en nitrates beaucoup moins toxiques et bénéfiques pour les plantes par des bactéries présentes naturellement ou artificiellement (plantées) dans l'aquarium.
Les bactéries Nitrosomonas transforment l'ammoniac en nitrites, eux-mêmes très toxiques pour les animaux aquatiques. Et les bactéries Nitrobacter transforment les nitrites en nitrates. Nitrates qui sont alors absorbés par les plantes présentes dans l'aquarium.
Lors du premier remplissage de l'aquarium, ces bactéries sont absentes de l'eau. Les populations de bactéries apparaissent progressivement à partir du moment où l'aquarium est rempli. Le lancement du cycle nécessite, selon le contexte, entre trois et huit semaines durant lesquelles l'eau de l'aquarium reste toxique. L'accumulation d'ammoniac provoque la prolifération de la première population de bactéries. Ces bactéries transforment l'ammoniac en nitrites. Au bout de quelques jours l'accumulation de nitrites due à la première population de bactéries provoque la prolifération de la deuxième population de bactéries, qui fait retomber la concentration de nitrites. La concentration de nitrites redescend jusqu'à être totalement indétectable. À partir de ce moment, l'aquarium est sain pour les poissons.
La plupart des aquariums ont un excédent de poissons, et pas assez de plantes par rapport à la quantité de nitrates produites. Les nitrates finissent par s'accumuler dans l'eau, et il est alors nécessaire de les diluer régulièrement au bout d'une à deux semaines en remplaçant une part de l'eau de l'aquarium par de l'eau « neuve ». La fréquence des remplacements dépendra de la sensibilité des animaux. De nombreux poissons supportent des concentrations jusqu'à 50 mg/L. Par contre, les coraux « durs » ne supportent pas des concentrations supérieures à 2 mg/L.
Les bactéries dégradant l'ammoniac et les nitrites sont présentes dans l'eau, le sol de l'aquarium et le filtre. Le brassage provoqué par le filtre et la pompe amène l'eau riche en oxygène de la surface vers le fond, et assure la survie des bactéries — qui ont besoin d'oxygène pour vivre. Il sert également à mettre en contact les bactéries présentes dans le filtre avec l'eau de l'aquarium[54].
À partir du moment où le cycle de l'azote est en route, on dit que l'aquarium est cyclé : l'ammoniac et les nitrites qui se forment dans l'eau sont immédiatement transformés, et les concentrations restent à des niveaux indétectables. Il peut cependant se retrouver intoxiqué accidentellement, si un excès de nourriture y est versé, ou si le cadavre d'un gros poisson y est laissé à se décomposer.
Ammonification
C'est la production d'ammonium ou d'ammoniac du fait d'une activité biologique, à partir soit de matière organique en décomposition, soit par la réduction de nitrate.
À partir de la matière organique
L'ammonification à partir de la matière organique a pour but de produire une source d'azote directement utilisable par les organismes pour leur propre synthèse[55].
Réalisée uniquement par une biomasse microbienne :
- bactéries gram- (Entérobactérie, Pseudomonacea, etc.) ;
- bactéries gram+ (Bacillus…) ;
- bactéries gram+ (Entérocoques…) ;
- champignons.
À partir des nitrates
Elle a pour buts :
- de produire de l'énergie par réduction des nitrates en ammonium[56] ;
- de produire une forme d'azote assimilable. C'est la dénitrification assimilatrice ou l'ammonification assimilatrice.
Application en traitement des eaux usées
Les stations d'épuration ont notamment comme fonction de dégrader les formes d'azote moléculaire et ammoniacal en nitrates, puis éventuellement de provoquer une dénitrification (ou dénitratation) afin de réduire la concentration de cet élément dans les eaux rejetées.
Nomenclature
Source[57].
- HNO3 : acide nitrique
- HNO2 : acide nitreux
- NO3− : ion nitrate
- NO2− : ion nitrite
- NH4+ : ion ammonium
- NH3 : ammoniac
- NH4OH : ammoniaque
- N2 : diazote
Illustrations
 Mesure/modélisation sur un an (2006-2007) des flux d'ammoniac en forêt et zone agricole de 500 ha à Lillington (Caroline du Nord), du pré-semis à la sénescence de la plante.
Mesure/modélisation sur un an (2006-2007) des flux d'ammoniac en forêt et zone agricole de 500 ha à Lillington (Caroline du Nord), du pré-semis à la sénescence de la plante.
Notes et références
- « L’Importance du cycle de l’azote », sur Blogue du bureau d’études PermacultureDesign (consulté le )
- Gordon W. Holtgrieve et al. (2011), "A Coherent Signature of Anthropogenic Nitrogen Deposition to Remote Watersheds of the Northern Hemisphere", Science Magazine, 16 décembre 2011, 1545-1548 (résumé)
- Electrons, life and the evolution of Earth's oxygen cycle. Paul G Falkowski, Linda V Godfrey. Philos Trans R Soc Lond B Biol Sci. 2008 Aug 27; 363(1504): 2705–2716.
- Smith, B., R. L. Richards, and W. E. Newton. 2004. Catalysts for nitrogen fixation : nitrogenases, relevant chemical models and commercial processes. Kluwer Academic Publishers, Dordrecht ; Boston.
- Feedbacks Between the Nitrogen, Carbon and Oxygen Cycles. Ilana Berman-Frank et al. In Nitrogen in the Marine Environment, 2008, Elsevier.
- Lee Chi Chung, Ribbe Markus W. et Hu Yilin, The Metal-Driven Biogeochemistry of Gaseous Compounds in the Environment, vol. 14, Springer, coll. « Metal Ions in Life Sciences », , 333 p. (ISBN 978-94-017-9268-4, PMID 25416393, DOI 10.1007/978-94-017-9269-1_6), « Chapter 7. Cleaving the N,N Triple Bond: The Transformation of Dinitrogen to Ammonia by Nitrogenases »
- Yvon R. Dommergues et François Mangenot, Écologie microbienne du sol, Masson et Cie, , p. 198.
- Robert Barbault, Écologie générale : Structure et fonctionnement de la biosphère, 5e édition, Dunod, p. 32
- (en) Robert K. Poole, Advances in microbial physiology, Academic Press, , p. 16.
- The Evolution and Future of Earth's Nitrogen Cycle. Donald E. Canfield et al., Science, 330, 192 (2010).
- A reduced, abiotic nitrogen cycle before the rise of oxygen. Ward, Lewis M, James Hemp, and Woodward W. Fischer. in Microbial Evolution and the Rise of Oxygen: the Roles of Contingency and Context in Shaping the Biosphere through Time. Thesis, Lewis Michael Ward, Caltech May 2017.
- J. M. Holloway et R. A. Dahlgren (2002), "Nitrogen in rock: occurrences and biogeochemical implications", Global Biogeochemical Cycles, 16(4), 65-1.
- C. Allègre, G. Manhés et E. Lewin (2001), Chemical composition of the Earth and the volatility control on planetary genetics, Earth Planet. Sci. Lett., 185, 46–69.
- W. H. Schlesinger, Biogeochemistry – An Analysis of Global Change, p. 588, Academic, San Diego, Calif., 1997.
- N. Story-Maskelyne, "On the mineral constituents of meteorites", Philos. Trans. R. Soc., Londres, 160, 189–214, 1870.
- B. Mason, "The Enstatite Chondrites", Geochim. Cosmochim. Acta, 30, 23–39, 1966.
- K. Keil, "Mineralogical and chemical relationships among enstatite chondrites", J. Geophys. Res., 73, 6945–6976, 1968
- G. E. Ericksen (1981) "Geology and origin of the Chilean nitrate deposits", U.S. Geol. Surv. Prof. Pap. 1188, 37,
- G. E. Ericksen, J. W. Hosterman et P. St. Amand, "Chemistry, mineralogy, and origin of the clay-hill nitrate deposits, Amargosa River valley, Death Valley region, California, U.S.A.", Chem. Geol., 67, 85–102, 1988.
- G. R. Mansfield et L. Boardman, "Nitrate deposits of the United States", U.S. Geol. Surv. Bull., 838, 107, 1932.
- C. A. Hill, "Mineralogy of Kartchner Caverns, Arizona", J. Cave Karst Stud., 61, 73–78, 1999
- J. E. J. Martini, "Sveite, a new mineral from Autana Cave, Territorio Federal Amazonas, Venezuela", Trans. Geol. Soc. S. Afr., 83, 239–241, 1980.
- M. E. Wallace, et A. Pring, "Gerhardtite, a copper hydroxy-nitrate from the Great Australia mine, Cloncurry, Queensland", Aust. Mineral., 5, 51–54, 1990.
- J. Toussaint, « Sur la gerhardtite de Likasi », Bull. Soc. Geol. Belgique, 79, 233–235, 1956.
- S. A. Williams, "Gerhardtite from the Daisy shaft, Mineral Hill mine, Pima County, Arizona", Ariz. Geol. Soc. Dig., 4, 123, 1961.
- E. Effenberger, "Likasite, Cu3(OH)5(NO3)(2H2O: Revision of the chemical formula and redetermination of the crystal structure", Neues Jahrbuch Mineral. Monatsh., 1986, 101–110, 1986.
- J. R. Keys, "Origin of crystalline, cold desert salts in the McMurdo region, Antarctica", Geochim. Cosmochim. Acta, 45, 2299–2306, 1981.
- G. E. Ericksen, J. J. Fahey et M. E. Mrose, "Humberstonite, Na7K3Mg2(SO4)(NO3)2(6H2O, a new saline mineral from the Atacama Desert, Chile", Geol. Soc. Am. Spec. Pap., 115, 59, 1968.
- J. K. Odum, P. L. Hauff et R. A. Farrow, "A new occurrence of ammoniojarosite in Buffalo, Wyoming", Can. Mineral., 20, 91–96, 1982
- Earl V. Shannon, "Ammoniojarosite, a new mineral of the jarosite group from Utah", American Mineralogist, 12, 424–426, 1927
- G. E. Dunning et J. F. Cooper Jr., "History and minerals of The Geysers, Sonoma County, California", Mineral. Rec., 24, 339–354, 1993.
- E. T. Allen et A. L. Day, "Steam wells and other thermal activity at The Geysers, California", Carnegie Inst. Washington Publ., 378, 42, 1927.
- E. Goldsmith, "On boussingaultite and other minerals from Sonoma County, California", Acad. Nat. Sci. Philadelphia, 28, 264–266, 1877.
- M. Q. Jan, P. S. B. Colback et M. Ahmad, "Low-temperature secondary minerals from Tarbela", Geol. Bull. Univ. Peshawar, 18, 189–197, 1985.
- G. M. Valentino, G. Cortecci, E. Franco, and D. Stanzione, "Chemical and isotopic compositions of minerals and waters from the Camp Flegrei volcanic system, Naples, Italy", J. Volcanol. Geotherm. Res., 91, 329–344, 1999.
- A. Parwel, R. Ryhage, and F. E. Wickman, "Natural variations in the relative abundances of the nitrogen isotopes", Geochim. Cosmochim. Acta, 11, 165–170, 1957.
- R. Martin, K. A. Rodgers, et P. R. L. Browne, "The nature and significance of sulphate-rich, aluminous efflorescences from the Te Kopia geothermal field, Taupo Volcanic Zone, New Zealand", Mineral. Mag., 63, 413–419, 1999.
- P. J. Bridge, et R. M. Clark, "Mundrabillaite - A new cave mineral from Western Australia", Mineral. Mag., 47, 80–81, 1983.
- L. C. Cavé, M. V. Fey, and D. K. Nordstrom, "Dissolution rate of apophyllite: The effects of pH and implications for underground water storage", in Proceedings of the 10th International Symposium on Water Rock Interaction, Villasimius, Sardinia, Italy, edited by R. Cidu, pp. 251–254, A. A. Balkema, Brookfield, Vt., 2001.
- H. M. Hori, T. Nagashima, K. Yamada et R. Miyawaki, Ammonioleucite, a new mineral from Tatarazawa, Fujioka, Japan, Am. Mineral., 71, 1022–1027, 1986
- S. P. Altaner, J. J. Fitzpatrick, M. D. Krohn, P. M. Bethke, D. O. Hayba, J. A. Goss et Z. A. Brown, Ammonium in alunites, Am. Mineral., 73, 145–152, 1988
- Richard C. Erd, D. E. White, J. J. Fahey et D. E. Lee, Buddingtonite, an ammonium feldspar with zeolitic water, Am. Mineral., 49, 831–850, 1964.
- M. D. Krohn, "Preliminary Description of a mineral-bound ammonium locality in the Cedar Mountains, Esmerelda County, Nevada", U.S. Geol. Surv. Open File Rep., 89-637. 12, 1989
- K. Kimbara et T. Nishimura, "Buddingtonite from the Toshichi Spa, Iwate Prefecture, Japan", J. Min. Soc. Jpn., 15, 207–216, 1982.
- R. A. Gulbrandsen, "Buddingtonite, ammonium feldspar, in the Phosphoria Formation, southeastern Idaho", J. Res. U.S. Geol. Surv., 2, 693–697, 1974.
- F. C. Loughnan, F. I. Roberts et A. W. Lindner, "Buddingtonite, NH4 feldspar in the Condor Oilshale Deposit, Queensland, Australia", Mineral. Mag., 47, 327–334, 1983.
- S. Higashi, "Tobelite, a new ammonium dioctahedral mica", Mineral J., 11, 16–27, 1982.
- P. N. Wilson, W. T. Parry et W. P. Nash, "Characterization of hydrothermal tobelitic veins from black shale, Oquirrh Mountains, Utah", Clays and Clay Minerals, 40, 405–420, 1992.
- V. A. Drits, H. Lindgreen et A. L. Salyn, "Determination of the content and distribution of fixed ammonium in illite-smectite by X-ray diffraction: Application to North Sea illite-smectite", Am. Mineral., 82, 79–87, 1997
- (en) Regina Temiño Boes, Anthropogenic alteration of the nitrogen cycle in coastal waters: Case studies from the Mediterranean Sea and the Gulf of Mexico, Universitat Politècnica de València, (lire en ligne).
- (en) R. Maranger, N. Caraco, J. Duhamel et M. Amyot, « Nitrogen transfer from sea to land via commercial fisheries », Nature Geoscience, vol. 1, , p. 111-112 (lire en ligne).
- Mississippi River Water Quality and the Clean Water Act: Progress, Challenges, and Opportunities
- Henry Steinberg, Greenpeace
- Hervé Widmer, « Ammoniac en aquarium marin », sur Mr Récif Captif, (consulté le )
- « Cycle de l'azote: humification, ammonification et nitrification », sur www.bio-enligne.com (consulté le )
- « Cycle de l'azote », sur www2.ggl.ulaval.ca (consulté le )
- D. Balou, E. Fabritius E. et A. Gilles, Toute la chimie 2e période, Ellipses, Paris, 2004
Voir aussi
Articles connexes
Liens externes
- Cycle de l'azote, schéma du cycle de l'azote en aquarium.
- Cycle de l'azote, explications sur la dégradation des composés azotés pour l'équilibre de l'aquarium.
Bibliographie
- James J. Elser, A World Awash with Nitrogen, Perspective Geochemistry Science, 2011-12-16, vol. 334, no 6062, p. 1504-1505, DOI:10.1126/science.1215567.
 Portail de la biochimie
Portail de la biochimie  Portail de l’agriculture et l’agronomie
Portail de l’agriculture et l’agronomie  Portail de l’aquariophilie
Portail de l’aquariophilie