Cyrtarachninae
Les Cyrtarachninae sont une sous-famille d'Araneidae réunissant aujourd'hui deux tribus : les Cyrtarachnini et les Mastophorini, cette dernière incluant les fameuses «Araignées à bolas» (araignées chassant leurs proies en faisant tournoyer un fil de soie lesté par des «perles» visqueuses).
Distribution géographique
Leur répartition est mondiale, du continent américain à l'Australie et l'Extrême Orient, en passant par l'Afrique et même l'Europe où une espèce, Cyrtarachne ixoides (ixodoides), les représente en France[1],[2].
Anatomie externe

Chez la plupart des espèces de Cyrtarachninae, l’abdomen des femelles adultes est proportionnellement plus large que le céphalothorax qu’il tend à recouvrir en le dissimulant plus ou moins et peut être très étalé dans le sens transversal (Pasilobus) (fig.1,D). Sa forme globale et son ornementation, motifs et fond de couleurs vives, peuvent simuler certains Insectes, par exemple des Coccinelles (Paraplectana coccinella, Paraplectana tsushimensis)(Fig.1,C). Il peut être également pourvu de tubercules ou verrucosités de tailles, formes, coloris variés (Pasilobus bufoninus, Cyrtarachne inaequalis, C.bufo, Poecilopachys australasia) (Fig.1,B,E). Le prosoma ou céphalothorax peut également présenter des excroissances en tubercules ou cornes (Mastophora), ces dernières pouvant simuler une rangée d'épines (Cladomelea debeeri) (Fig.2).

Cette morphologie particulière peut favoriser le mimétisme de l'araignée avec son environnement (mimétisme batésien), en étant par exemple associé à une couleur d’ensemble jaune-verdâtre fondant l' araignée dans la végétation (Taczanowskia)(Fig.1,G), ou en permettant une ressemblance (rare) avec des insectes (Fig.1,C), avec des débris végétaux ou des excréments d’oiseaux tombés sur la végétation (Celaenia, Mastophora extraordinaria: «Bird-dropping spiders» des anglo-saxons)(Fig.1,H), ou encore avec la coquille de mollusques terrestres (Cyrtarachne conica, Cyrtarachne inaequalis, Cyrtarachne bufo)(Fig.1,B).

Par ailleurs, la morphologie générale et la coloration peuvent prêter à confusion avec les Gasteracanthes, comme dans le cas d’Aranoethra cambridgei dont le jaune éclatant (Fig.1,L) rappelle celui de Gasteracantha remifera. Emerit[1]a d’ailleurs tenté un rapprochement phylogénique audacieux entre les Cyrtarachninae, surtout mâles immatures de Cyrtarachne "ixodoides", et les Gastéracanthes, lien basé sur la sigillotaxie (plaques chitinisées) de l’abdomen et ses tubercules.
Les mâles, du moins chez les Mastophorini, sont sensiblement plus petits que les femelles (Fig.3). Il existe donc alors un dimorphisme sexuel volumétrique marqué comme chez les Argiopes et les Néphila.

Industrie séricigène: les toiles et les modes de capture des proies
Les Araignées de la sous-famille des Cyrtarachninae construisent des toiles d’une extraordinaire diversité structurale (Stowe,1986)[3] opposant les deux tribus constitutives de la sous-famille. Dans le cadre de cette industrie séricigène entre le tissage de cocons ovigères globuleux (Mastophora, Celaenia)(Fig.4) ou fusiformes (Cyrtarachne ixoides, Dicrostichus magnificus)(Fig.5).

Selon une première hypothèse (Robinson & Robinson, 1975[4]; Eberhard1980[5]), les toiles auraient évolué par réduction progressive en trois étapes qui se sont succédé au fil du temps: le type à fils en pont («spanning-thread webs» des anglo-saxons), le type en triangle, le type à bolas et, en fin de compte, la disparition pure et simple de tout édifice soyeux (Scharff & Coddington, 1997)[6].
Elle a été infirmée plus récemment par l' étude de phylogénie moléculaire ardue de Tanikawa et al. (2014)[7], puis celle de Scharff et al. (2020)[8].
Tribu des Cyrtarachnini
Les genres Cyrtarachne, Pasilobus, Poecilopachys et Paraplectana tissent des toiles orbiculaires très modifiées car pourvues d’un petit nombre de radii et de fils visqueux largement espacés à disposition non spiralée, nommés en anglais ‘spanning-threads”, soit “fils en pont”, (Clyne, 1973[9]; Chigira,1978[10]; Robinson, 1980[11]; Shinkai, 1992). Leur construction « laborieuse » a été décrite en détails par Stowe (1986)[3], d’après différents auteurs dont s’inspirent également les dessins joints.
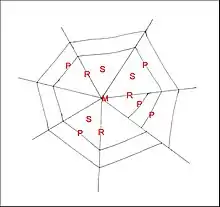
La toile des Cyrtarachne exotiques se compose d’une charpente de forme variable, en triangle ou en polygone plus complexe, de radii divergeant à partir d’un moyeu subcentral et de fils en pont pouvant former des cercles complets au travers des secteurs (Fig.6).
Elle peut atteindre 1 m de diamètre.
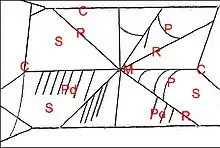
La toile de notre espèce indigène Cyrtarachne ixoides , étudiée en détail par Olin (2013) dans les Landes se rencontre souvent entre deux buissons ou herbes hautes, à une hauteur de 1 à 2 m, plus rarement jusqu’à 3 ou 4 m dans un arbre. Cet édifice, mieux visible quand il est emperlé de rosée matinale, se compose d’une charpente en cadre rectangulaire, s’attachant typiquement à quatre supports, et d’un ensemble de 8 à 10 radii convergeant vers un moyeu (Fig.7,8)

renforcé de soie plus dense et, dans les secteurs, de fils en pont très gluants, sans disposition spiralée. Ces fils de capture forment, de profil, des arceaux sous la toile lorsqu’ils sont rattachés à deux rayons successifs par leurs extrémités ou pendent verticalement, lorsque l’une de ces dernières s’est rompue, spontanément ou sous l’impact d’une proie volante (Fig.7).
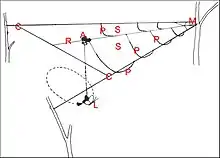
La toile de Pasilobus, longue d’environ 50 cm, a une charpente triangulaire, un moyeu totalement excentré à l’apex, un unique radius axial et, de ce fait, seulement deux secteurs traversés par quelques fils en pont (Fig.8).L’ Araignée Pasilobus sp., étudiée en Nouvelle Guinée (Morobe District) par les Robinson (Robinson & Robinson, 1975)[12], construit cet édifice de nuit sur les arbustes et les petits arbres. Plus ou moins horizontal, cet édifice a une charpente triangulaire divisée en deux moitiés par un fil médian tendu de l’ angle apical au milieu de la base, ainsi divisée. De ce fil axial pendent de 4 à 11paires de fils en “pont” largement espacés. Seuls constituants adhésifs de la toile, ils sont visqueux dans une partie seulement de leur longueur, solidement fixés au seul niveau de leur jonction avec le fil médian mais, en revanche, sont facilement rompus à leur insertion sur le fil de charpente latéral ("low‐shear joint"). La toile de Paraplectana, telle que P. tsushimensis (Chigira,1978), d’un diamètre moyen de 70 cm, a une charpente plus ou moins incomplète, un moyeu très excentré, 4 à 7 radii et des fils en pont dans les secteurs plus ou moins ouverts qu’ils délimitent (Fig.9).
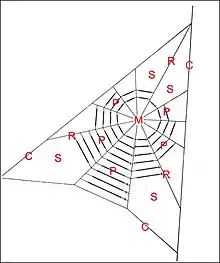
La toile de Poecilopachys australasia (Clyne,1973), d’environ 80 cm de diamètre, montre également une charpente triangulaire, mais il s’y inscrit une ébauche d’orbe à moyeu excentrique, 7 à 12 radii, une dizaine de secteurs et jusqu’à un maximum de 13 fils en pont dans ces derniers (Fig.10).
Lorsqu’un insecte vient à heurter un fil en pont de ces toiles, l’une des extrémités de ce fil, dite «joint de cisaillement» (“low-shear joint”) se libère du radius, tandis que l’autre lui reste fermement attachée (Cyrtarachne Arita, 1963; Poecilopachys Clyne, 1973; Paraplectana Chigira, 1978; Pasilobus Robinson, 1980; Cyrtarachne Shinkai, 1992)(Fig.8). Cette caractéristique différencie nettement les fils en pont des spires de toiles orbiculaires habituelles qui sont attachés fermement aux radii et ne s’en détachent point par rupture. Le caractère gluant du fil en pont est lié aux gouttelettes visqueuses qui y sont fixées et retiennent la proie jusqu’à ce que l’ araignée la hale vers elle (Fig.4).
Dans le cas de Pasilobus, lorsqu’un Insecte heurte un «pont» en volant(Fig. 8), ce dernier se rompt et pend au-dessous de la toile, attachant ainsi au fil médian la proie qui continue à voler en tournoyant et finit par s’immobiliser. L’araignée se précipite vers l’attache le long du fil axial, hale le fil en “pont” et mord enfin l’ insecte englué sur ce dernier.(Robinson)[4].
Tribu des Mastophorini : araignées à bolas
Apparemment bien différente est l’industrie séricigène des genres Mastophora (Amérique), Cladomelea et Acantharachne (Afrique), Ordgarius ou Dicrostichus (Australie) dont la toile captrice est réduite à un « bola », fil terminé ou garni par une ou plusieurs masses visqueuses alors superposées que l’ araignée femelle fait tourner au bout de l’une de ses pattes lorsque s’approche, de nuit, une proie volante (Eberhard, 1980[5]; Shinkai and Shinkai, 2002[13]; Blackledge et al., 2011[14]). Chacun de ces genres diffère des autres par sa tactique de manipulation du bola, tenu avec une patte qui le projette vers la proie parvenue à portée : patte I chez Mastophora et Agatostichus, patte II chez Dicrostichus (Longman1922 ; Mac.Keown,1952) avec tournoiement à l'approche de la victime et patte III chez Cladomelea où la rotation est permanente (Akerman, 1923).
Tribu des Mastophorini : araignées sans bolas
A l’extrême, les Araignées femelles de deux autres genres de la même tribu, Celaenia et Taczanowskia (Amérique) ne tissent aucun piège et capturent directement avec leurs pattes étendues (McKeown, 1952; Eberhard, 1981; Forster and Forster, 1999).
Points communs

Suspension de la proie
En fait, bien que les structures des pièges diffèrent grandement, il existe des similitudes frappantes entre une proie suspendue à un fil en pont rompu et ballant et celle qui a été prise à la « volée » avec un bola.
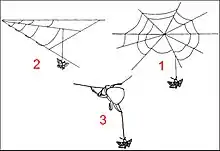
Adhésivité
L’adhésivité des gouttelettes de fils en pont et du globule de bola est extrême. Cependant, chez Cyrtarachne, elle décroîtrait en quelques heures jusqu’à devenir nulle (Cartan et Miyashita, 2000)[15], sauf dans des conditions de forte humidité où elle reste élevée (Baba et al., 2014)[16]. Il en serait de même pour les “énormes” globules de bolas qui diminuent très sensiblement de taille en seulement 90 minutes (Eberhard, 1980)[5], réduction d’adhésivité non observée dans les toiles orbiculaires des autres Araneidae.
Attraction des proies par mimétisme chimique agressif phéromonal
Les “Bolas spiders” ont un autre caractère unique dans le fait que leurs femelles adultes attirent exclusivement des Lépidoptères nocturnes, tous mâles, en émettant une substance chimique volatile véhiculée par l’air (Stowe et al., 1987;Yeargan, 1994)[17],[18] selon un mécanisme connu comme mimétisme sexuel phéromonal. En revanche, les mâles et les juvéniles n’attirent pas des Lépidoptères mais des Diptères Psychodidae, également mâles (Yeargan and Quate,1996,1997)[19],[20].
A l’extrême, les Araignées femelles de deux autres genres de la même sous-famille, Celaenia et Taczanowskia (Amérique) ne tissent aucun piège et capturent directement leurs proies, toujours des Lépidoptères nocturnes mâles (Stowe, 1986)[3], avec leurs pattes étendues (McKeown, 1952; Eberhard, 1981; Forster and Forster, 1999). Ici encore, un mimétisme chimique agressif phéromonal doit être utilisé par ces Araignées, d’autant plus que, lorsqu’elles sont jeunes, leurs proies consistent aussi en Psychodidae (Forster and Forster, 1999).
Support anatomique
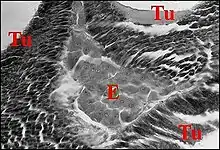
La structure ou, à l'inverse, l'absence de piège, et l'attraction des proies trouvent une explication logique révélée par la seule histologie de l'opisthosoma, débité en coupes sériées (C.H) respectivement dans les glandes séricigènes et dans l'existence d'un organe particulier remarquable, le tissu folliculaire endocrinoïde abdominal (Fig.12,13), tel que Lopez l'a décrit pour la première fois chez les Mastophora.[21]
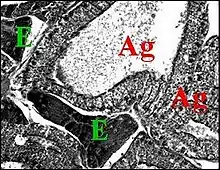
Il l'a retrouvé ensuite, toujours par étude histologique chez Celaenia, membre des Mastophorini (Fig.14), et Poecilopachys, une représentante des Cyrtarachnini (Fig.15)[22].


Le même auteur (1998) a montré que les glandes à soie agrégées sont absentes lorsque l'Araignée Mastophorine Celaenia qui ne construit aucune toile et n'élabore pas de glue. L'absence de cette catégorie de glandes séricigène est d'ailleurs une caractéristique d'autres Araneidae (Cyrtophora, Mecynogea)[23] dont la toile de capture est dépourvue de tout matériel gluant dans son orbe modifiée.
En revanche, les agrégées sont particulièrement bien développées dans les genres (Mastophora,Poecilopachys) qui produisent quantité de matériel gluant (Fig.12,13,15).
Histoire phylogénétique
Elle est ardue et a débuté avec le travail original de Simon (1892)[24].
Les Cyrtarachninae constituent un groupe sous-familial d’ araignées dans la famille des Araneidae (Araignées dites «à toile orbiculaire»). Ce groupe a été traité de différentes manières, au prix de remaniements oiseux.
Il est apparu initialement sous le nom de Cyrtarachneae (Simon, 1892)[24] incluant 5 genres, à savoir Paraplectana, Aranoethra, Pasilobus, Cyrtarachne et Poecilopachys. En cette occasion, Simon créa aussi le groupe des Glyptocranieae, incluant les genres Agatostichus (reconnu encore par Gertsch en 1955 mais aujourd’hui rattaché à Mastophora), Glyptocranium (traité de même), Cladomelea et Ordgarius (ex Dicrostichus)(Eberhard,1982). De même, il plaça deux genres, Celaenia and Taczanowskia dans le groupe des Celaenieae. Les trois groupes ainsi créés, Cyrtarachneae, Mastophoreae, Celaenieae, furent traités comme des tribus, à savoir les Cyrtarachnini, Mastophorini and Celaeniini.
Les dénominations ont ensuite varié, la plus large, celle des Cyrtarachninae sensu lato (s.l.), renfermait les trois groupes originaux de Simon[24], y compris, bien qu’à priori incongrues dans cette place, les araignées à bolas (tribu des Mastophoreae de Mello-Leitão,1931)[25]. Emerit (1978) considéra à tort les deux premiers Excavata comme des sous-familles, les Cyrtarachninae et les Mastophorinae. Plus tard, Eberhard (1980, 1982)[5],[26]dans des études comportementales montra que les trois groupes de Simon partageaient bien des caractéristiques communes et pouvaient être réunis.
Plus récemment (1997), une analyse phylogénétique de la famille des Araneidae a été publiée par Scharff & Coddington[6]. Ces auteurs ont inclus les divers genres de deux des groupes de Simon, Cyrtarachneae et Mastophoreae, les considérant comme monophylétiques, donc étroitement apparentés («sisters») et les combinant dans la sous-famille des Cyrtarachninae, en excluant toutefois la totalité des Celaenieae. Ils soulignent que la sous-famille a une biologie assez cohérente, une tendance marquée à la réduction de la toile et une évolution compensatrice du mimétisme chimique agressif.
Plus près de nous encore, Tanikawa et al. (2014)[7] par une étude de phylogénie moleculaire ardue, puis Scharff & a. (2020)[8] ont infirmé cette conception qui ne s’accorde pas avec les relations évolutives déduites antérieurement. D’une part, les Araignées à toiles triangulaires (Pasilobus) doivent rester incluses dans le groupe confectionnant des “ spanning-thread webs », sous l’étiquette de « clade C ». D’autre part, les espèces utilisant des bolas et celles qui ne construisent aucun piège forment un groupe monophylétique complètement séparé du précédent en tant que « clade B » descendant d’un ancêtre commun (Tanikawa et al.).
Il se pourrait enfin que, selon une conception de A.Lopez, à confirmer ou non par des recherches histologiques ultérieures, seules valables en ce genre de détection, il existe un lien anatomique original (synapomorphie), caractère dérivé, entre tous les représentants des Cyrtarachninae: l'existence d'un tissu endocrinoïde abdominal, démontrée dans deux genres de Mastophorini (Mastophora, Celaenia)(Fig.14) et jusqu'ici un seul (Poecilopachys) chez les Cyrtarachnini (Fig.15) à explorer globalement sur le plan de l'anatomie interne.
Biogéographie
La richesse en Araignées à bolas est la plus élevée dans le Nouveau Monde, avec 51 espèces de Mastophora, incluant Agatostichus , contre une seule espèce connue de Cyrtarachne.
En Afrique, les Araignées à bolas ne sont représentées que par les genres Cladomelea et Acantharachne.
En Asie sensu lato, il n’y aurait en revanche que 11 espèces du genre australien Ordgarius ou Dicrostichus contrastant avec la richesse en « "spanning-thread web spiders» connues seulement (Platnick, 2014) dans cette partie du monde. Parmi elles, 43 espèces de Cyrtarachne.
Cela suggère une origine américaine du clade à bolas ou sans toile, et une origine asiatique du clade à fils en pont («spanning threads»). La diversification de l’ ancêtre commun en ces deux clades peut être survenue en association avec leur division géographique.
Les détails de la phylogéographie restent à préciser.
Bibliographie
Eberhard, WG,1980 - The natural history and behavior of the bolas spider Mastophora dizzydeani sp. n. (Araneidae). Psyche 87, p.143–169.
Eberhard ,WG., 1981 - Notes on the natural history of Tackzanowskia sp. (Araneae: Araneidae). Bull Br Arachnol Soc., 5, p.189–204
Lopez,A.avec M.K.Stowe et J.C.Bonaric, 1985 - Anatomie interne de l’ «Araignée à bolas» nord-américaine Mastophora cornigera (Hentz, 185o) (Araneae: Araneidae) après sa sortie du cocon. Pub.Scient.accélérées (Rémy Chauvin ed.), n° 8, p. 1–9.
Lopez,A.avec L. Juberthie-Jupeau et M.K.Stowe, 1986 – L’appareil séricigène de Kaira alba (Hentz). Mem.Soc.roy. belge d’Entomologie, 33, p. 119-128.
Lopez,A., 1998.- L’origine des odeurs attractives de Papillons et de Diptères mâles chez certaines Araignées exotiques (genres Kaira, Mastophora et Celaenia: Araneidae): une approche histologique. Bull. Soc.Et.Sci.nat. Béziers, 17, 58, p.9-24.
Simon,E.,1892 - Histoire naturelle des araignées, (2nd ed.), Roret, Paris, 1, p.876-895.
Shinkai, A. & E Shinkai, 2002 - The natural history, bolas construction,and hunting behavior of the bolas spider, Ordgarius sexspinosus (Thorell) (Araneae: Araneidae). Acta Arachnol 51, p. 149–154
Tanikawa,A, Shinkai,A., & T. Miyashita, 2014 - Molecular Phylogeny of Moth-Specialized Spider Sub-Family Cyrtarachninae, which Includes Bolas Spiders. Zoological Science 31(11), p.716-720.
Robinson M.H. & B.Robinson 1975 - Evolution beyond the orb web: The web of the areneid spider Pasilobus sp., its structure, operation and construction. Zool J Linn Soc 56, p. 301–314.
Stowe, M.,1986 - Prey specialization in Araneidae. In “Spiders –Webs, Behaviour, and Evolution” Ed by WA Shear. Stanford Univ Press, Stanford, p. 101–132
Clyne,D.,1973 - Notes on the web of Poecylopachys australasia (Griffith & Pidgeon)(Araneida : Argiopidae). Aust.Entomol.Mag.,I, p.23-29.
Robinson, MH.,1980 - The ecology and behavior of tropical spiders.Proc. Int .Arachnol. Congr., 8, p.13–32.
Chigira,Y. (1978) A note of the web of Paraplectana tsushimensis.Atypus, 72, p.19–24
Notes & références
- Emerit, M., « Cyrtarachne ixodoides, une araignée rare du Midi de la France : la pointe de pénétration d’une sous-famille tropicale. », Revue arachnologique série 1, 1, p. 23-31.,
- Odin, G.S., « Détection de toiles de Theridula sp. (Araneae, Theridiidae) et Cyrtarachne ixoides (Araneae, Araneidae), près de Dax (Landes). », Revue arachnologique, série 2, n° 4, juin 2017, p. 46-53.,
- (en) Stowe, M., « Prey specialization in Araneidae. », In “Spiders –Webs, Behaviour, and Evolution” Ed by WA Shear. Stanford Univ Press, Stanford, p. 101–132,
- (en) Robinson M.H. & B.Robinson, « Evolution beyond the orb web: The web of the areneid spider Pasilobus sp., its structure, operation and construction. », Zool J Linn Soc 56, p. 301–314.,
- (en) Eberhard, WG., « The natural history and behavior of the bolas spider Mastophora dizzydeani sp. n. (Araneidae). », Psyche 87, p.143–169.,
- (en) Scharff, N. & J.A. Coddington, « A phylogenetic analysis of the orb-weaving spider family Araneidae (Arachnida, Araneae). », Zool. J.Linn. Soc., 120, p. 355–434.,
- (ja) Tanikawa,A, Shinkai,A., & T. Miyashita, « Molecular Phylogeny of Moth-Specialized Spider Sub-Family Cyrtarachninae, which Includes Bolas Spiders. », Zoological Science 31(11), p.716-720.,
- (en) Scharff, N., Coddington, J.A., Blackledge, Todd, A., Agnarsson, I., Volker, W.,Szűts, T., Cheryl, Y. & Dimitrov Dimitar, « Phylogeny of the orb‐weaving spider family Araneidae (Araneae: Araneoidea), », Cladistics, 36, p.1–21.,
- (en) Clyne,D., « Notes on the web of Poecylopachys australasia (Griffith & Pidgeon)(Araneida : Argiopidae). », Aust.Entomol.Mag.,I, p.23-29.,
- (ja) Chigira,Y., « A note of the web of Paraplectana tsushimensis. », Atypus, 72, p.19–24,
- (en) Robinson, MH., « The ecology and behavior of tropical spiders.Proc. Int .Arachnol. Congr., 8, p.13–32. », Proc. Int .Arachnol. Congr., 8, p.13–32.,
- (en) Robinson M.H. & B.Robinson, « Evolution beyond the orb web: The web of the areneid spider Pasilobus sp., its structure, operation and construction. », Zool J Linn Soc 56, p. 301–314,
- (ja) Shinkai, A. & E. Shinkai, « The natural history, bolas construction,and hunting behavior of the bolas spider, Ordgarius sexspinosus (Thorell) (Araneae: Araneidae). », Acta Arachnol 51, p. 149–154,
- (en) Blackledge, TA, Kuntner, & M. Agnarsson, « The Form and Function of Spider Orb Webs: Evolution from Silk to Ecosystems. », In Casas, J. (ed.) Advances in Insect Physiology, Vol. 41, Academic Press, Burlington, p.175–262,
- (en) Cartan, CK & T.Miyashita, « Extraordinary web and silk properties of Cyrtarachne (Araneae, Araneidae): a possible link between orb-webs and bolas. », Biol J Linn Soc 71, p.219–235.,
- (en) Baba, YG, Kusahara, M, Maezono, Y & T. Miyashita, « Adjustment of web-building initiation to high humidity: a constraint by humidity-dependent thread stickiness in the spider Cyrtarachne. », Naturwissenschaften 101, p. 587–593,
- (en) Stowe M, Tumlinson JH. & RR.Heath, « Chemical mimicry: Bolas spiders emit components of moth prey species sex pheromones. », Science 236, p.964–967.,
- (en) Yeargan KV., « Biology of bolas spiders. », Annu. Rev. Entmol., 39, p. 81–99.,
- (en) Yeargan KV. & L.W.Quate, « Juvenile bolas spiders attract psychodid flies. », Oecologia, 106, p. 266–271.,
- (en) Yeargan K. V. & L.W. Quate, « Adult male bolas spiders retain juvenile hunting tactics. », Oecologia, 112, 4, p. 572-576.,
- Lopez,A., M.K.Stowe et J.C.Bonaric, « Anatomie interne de l’ «Araignée à bolas» nord-américaine Mastophora cornigera (Hentz, 185o) (Araneae: Araneidae) après sa sortie du cocon. », Pub.Scient.accélérées (Rémy Chauvin ed.), n° 8, p. 1–9.,
- Lopez,A., « L’origine des odeurs attractives de Papillons et de Diptères mâles chez certaines Araignées exotiques (genres Kaira, Mastophora et Celaenia: Araneidae): une approche histologique. », Bull. Soc.Et.Sci.nat. Béziers, 17, 58, p.9-24.,
- Lopez,A. & J. Kovoor, « Anatomie et histologie des glandes séricigènes des Cyrtophora (Araneae, Araneidae) : affinités et corrélations avec la structure et la composition de la toile. », Revue Arachnologique, vol. 4, p. 1-21.,
- Simon,E., « Histoire naturelle des araignées », Histoire naturelle des araignées(2nd ed.), Roret, Paris, 1, p.876-895.,
- (pt) Mello-Leitão, CF de, « Contribuicao ao estudo da tribu Mastophoreas. », Anais da Academia Brasileira de Ciências . 3, p. 65–74,
- (en) Eberhard ,WG., « Notes on the natural history of Tackzanowskia sp. (Araneae: Araneidae). », Bull Br Arachnol Soc., 5, p.189–204,
 Portail de l’arachnologie
Portail de l’arachnologie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire