Dictyostelium discoideum
Dictyostelium discoideum est une espèce de mycétozoaires. Cette amibe vivant sur les tapis de feuilles mortes dans les forêts, se nourrissant de bactéries et de levures, est utilisée comme organisme modèle de laboratoire. Elle est également appelée « amibe sociale ».
| Règne | Protozoa |
|---|---|
| Division | Dictyosteliomycota |
| Classe | Dictyosteliomycetes |
| Ordre | Dictyosteliales |
| Famille | Dictyosteliaceae |
| Genre | Dictyostelium |
Les études effectuées sur cet organisme ont permis de décrire de nombreuses voies de signalisation communes aux eucaryotes. Un autre domaine d'étude est celui de la chimiotaxie dont les mécanismes décrits peuvent être extrapolés à de nombreuses cellules, notamment des cellules humaines telles que les leucocytes, les fibroblastes, les cellules embryonnaires ou encore certaines cellules cancéreuses.
Cycle de vie
Chaque amibe unicellulaire est issue d'une spore. Ces amibes peuvent emprunter trois voies différentes. La première consiste en la transformation vers une forme de résistance appelée kyste. Ce kyste redonne une amibe quand les conditions redeviennent favorables. La seconde voie est celle de la reproduction sexuée qui comprend une étape d'agrégation entre plusieurs amibes pour donner un macrokyste qui après méiose donnera naissance à de nouvelles amibes. La troisième voie, la plus courante, est choisie par les amibes en cas de carence nutritionnelle. Dans ces conditions, Les amibes sécrètent un chimioattractant qui dans le cas de Dictyostelium discoideum est l'AMPc. L'agrégation de quelques amibes entraine la formation d'une forte source d'AMPc qui attire les amibes alentour. Après cette phase d'agrégation, les amibes forment un pseudo-plasmode. Ce pseudo-plasmode ressemble à une petite limace, pouvant atteindre quelques millimètres, constitué de milliers d'amibes agglomérées et vivant en société. Ce pseudo-plasmode peut persister pendant plusieurs jours à la recherche de conditions favorables. Il se forme ensuite un sporocarpe qui est la fructification du pseudo-plasmode. Ce sporocarpe est constitué d'une tige supportant une boule d'amibes qui se différencient en spores pour se disséminer.
Plusieurs étapes de ce cycle ont un intérêt fondamental en recherche:
- La chimiotaxie en réponse à l'AMPc.
- Le passage vers une forme de multicellularité.
- Le développement du sporocarpe.
Développement du sporocarpe
Lors de ce développement, certaines amibes constitueront la tige et mourront sans perpétuer leur patrimoine génétique alors que les autres amibes donneront des spores et pourront se disperser. On rapporte qu'environ 80% des cellules deviendront des spores alors que les 20% restants se dirigeront dans la tige et y subiront une mort cellulaire programmée[2]. Il existe des souches d'amibes tricheuses qui ne participent pas à la formation de la tige afin de se disperser. Ce phénomène de tricherie implique au moins une centaine de gènes qui commencent à être bien caractérisés.
Agriculture primitive
Des scientifiques américains ont montré que D. discoïdeum est capable de "cultiver" des bactéries[3]. En effet, ils ont constaté que les sporocarpes de certaines colonies (environ un tiers des échantillons naturels) contiennent des bactéries vivantes. Ces bactéries sont ensuite dispersées avec les spores, fournissant ainsi à l'amibe de la nourriture quel que soit l'endroit où elle se trouve. Cette association entre les amibes et les bactéries est stable, c'est-à-dire qu'une colonie capable de conserver des bactéries garde cette capacité à la génération suivante ; inversement, les amibes ne cultivant pas de bactéries ne le feront jamais.
Génome
Le génome de Dictyostelium discoideum a été séquencé. Sa taille est de 34 Mb répartis sur 6 chromosomes, il comporte également un chromosome palindromique codant l'ADNr. Ce génome est très riche en bases AT (~80 %) ainsi qu'en éléments répétés, rendant son analyse difficile.
Génétique
Des outils génétiques ont été développés. Il est possible de transformer Dictyostelium discoideum avec des plasmides intégratifs ou réplicatifs. La technique appelée REMI permet également de caractériser les mutants.
Multicellularité
D. discoideum, en tant qu’organisme modèle, occupe une place importante en recherche. C’est qu’il s’agit d’une amibe sociale, ce qui veut dire qu’elle se situe dans un flou entre la vie unicellulaire et la vie multicellulaire. Il s’agit donc de l’une des formes de vie les plus simples chez laquelle on peut observer plusieurs phénomènes retrouvés chez les organismes multicellulaires. En effet, comme les différents individus D. discoideum doivent former une tige et des spores pour la dispersion, ils doivent utiliser plusieurs procédés développementaux comme la signalisation cellulaire, l’apoptose, la différenciation cellulaire et l’adhésion intercellulaire.
Signalisation cellulaire
Les études sur les eucaryotes multicellulaires, comme les plantes et les animaux, ont montré que les dérivés réactifs de l’oxygène, comme le peroxyde d’hydrogène et les anions superoxydes, jouaient un rôle important dans la signalisation cellulaire, par exemple lors de la division cellulaire et de la différentiation cellulaire.
Les eucaryotes unicellulaires comme D. discoideum ont également des processus de signalisation réagissant aux dérivés réactifs de l’oxygène, mais de façon générale, ils n’en produisent pas activement comme chez les eucaryotes multicellulaires. Comme D. discodeum se trouve à l’embranchement entre la vie unicellulaire et la vie multicellulaire, des chercheurs, Bloomfield et Pears[4], se sont penchés sur l’utilisation de ces réactifs chez cette espèce afin de déterminer si la production active de dérivés réactifs de l’oxygène pouvait être apparue chez des organismes comme Dyctiostelium.
L’étude a montré que lors de carence nutritionnelle, donc lors du développement multicellulaire pour former la tige et les spores chez l’amibe, la production de superoxydes augmentait. Il a également été montré que lorsque la superoxyde dismutase, une enzyme inhibant la formation de la tige, était elle-même inhibée, la production de superoxydes était encore plus grande. Il a également été montré que les superoxydes étaient générés majoritairement au niveau de la membrane plasmique, bien que la respiration cellulaire en soit une source également. Finalement, des cellules de D.discoideum ont été modifiées génétiquement pour produire plus de superoxyde dismutase (et donc moins de superoxydes). Celles-ci étaient beaucoup moins efficaces pour former les agrégations nécessaires au développement multicellulaire de l’organisme.
Cette étude a donc permis de montrer que les superoxides jouent bel et bien un rôle important lors du début du développement de D.discoideum, notamment pour la signalisation cellulaire permettant l’agrégation. L’étude a aussi permis de savoir que ces superoxides sont principalement trouvés au niveau de la membrane cellulaire, ce qui indiquerait une enzyme oxydase NADPH comme source probable des superoxydes. Une enzyme similaire est également retrouvée chez les mammifères et les plantes, des eucaryotes multi-cellulaires. Comme ce type de signalisation cellulaire – l’utilisation de superoxydes produits par une enzyme oxydase NADPH – est également retrouvé dans les processus développementaux d’autres organismes, par exemple le développement de l’œil chez la drosophile ou des cellules neurales chez les mammifères, il est probable que l’évolution de cette signalisation cellulaire est directement lié avec l’évolution de la multicellularité.
Mort cellulaire programmée
Chez les organismes multicellulaires, la mort cellulaire programmée est utilisée dans le cadre du développement et du fonctionnement habituel de l’organisme (homéostasie). L’apoptose est, chez ces organismes, la forme la plus fréquente de mort cellulaire programmée. Ce type de processus ne semblerait toutefois pas réservé aux multicellulaires. En effet, la différenciation des cellules de D. discoideum en cellules de tige s’apparente à une mort cellulaire programmée. De plus, même au cas où l’agrégation (et donc la différenciation) est impossible, des chercheurs (Tatischeff et al.)[2] ont observé une autre forme de mort cellulaire programmée ressemblant beaucoup à l’apoptose des mammifères. Ces chercheurs ont donc observé des cellules de D. discoideum dans des situations où l’agrégation était impossible mais où il y avait tout de même carence en nutriments (la situation originale débutant le processus de développement multicellulaire chez D. discoideum).
L’induction de la mort cellulaire chez les cellules était accélérée lorsqu’on les empêchait de s’agréger. La nature de cette mort a été observée en détail, révélant que la cellule changeait d’apparence pendant cette mort dans un ordre successif. De façon successive, les cellules de D. discoideum mourant de cette façon voyaient leur membrane plasmique se déformer, la production de blebs (bulles formées par la déformation du cytosquelette), perte de corps cellulaires apparente, réduction de la taille de la cellule et perte de réfringence. Cette succession de changements morphologiques ressemblent beaucoup aux processus que subissent les cellules lors de l’apoptose chez les organismes multicellulaires.
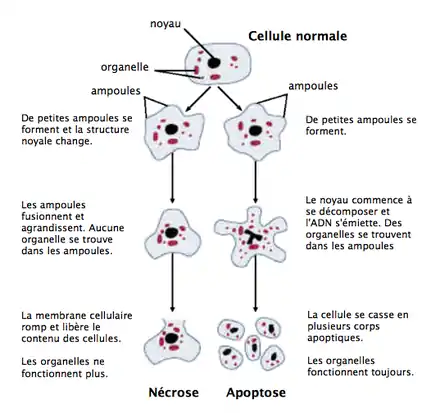
Une observation encore plus élaborée de ces cellules a permis de voir les processus intracellulaires responsables de ces changements morphologiques chez D. discoideum. Ces processus se sont révélés à être la formation de vacuoles, une digestion autophagique avant la dégradation de la membrane plasmique et une phagocytose des cellules mortes par ses voisines. Ces processus sont, encore une fois, des caractéristiques de l’apoptose chez les organismes multicellulaires comme les mammifères.
Finalement, l’étude a également révélé une perte de potentiel transmembranaire (donc une perméabilisation de la membrane) des mitochondries et une exposition accrue de phosphatidylsérine sur la partie externe de la membrane plasmique chez les cellules de D. discoideum mourantes. Ces deux observations sont également des conséquences de l’apoptose chez les mammifères.
En bref, les cellules de D. discoideum semblent avoir deux types de mort différentes : l’une lorsqu’elle peuvent s’agréger et former une tige composée de cellules mortes, et une autre lorsque l’agrégation est impossible. Dans le deuxième cas, les individus unicellulaires subissent un processus ressemblant beaucoup à l’apoptose des mammifères : changements morphologiques, autophagie très efficace, reconnaissance et élimination des cellules mourantes par phagocytose, baisse du potentiel transmembranaire mitochondrial et exposition accrue de phosphatidylsérine sur la membrane cellulaire. Cette découverte, la similarité de la mort cellulaire de l’amibe et de l’apoptose des mammifères, est importante puisqu’elle peut nous indiquer comment l’apoptose, un processus important dans le développement multicellulaire, a pu évolué en premier lieu chez des organismes unicellulaires comme D. discoideum. De plus, cette ressemblance nous permet de mieux comprendre l’apoptose en étudiant l’organisme modèle D. discoideum, et donc de transmettre ces connaissances dans le domaine biomédical.
Notes et références
- V. Robert, G. Stegehuis and J. Stalpers. 2005. The MycoBank engine and related databases. https://www.mycobank.org/, consulté le 23 avril 2019
- Irène Tatischeff, Patrice Xavier Petit, Alain Grodet et Jean-Pierre Tissier, « Inhibition of multicellular development switches cell death of Dictyostelium discoideum towards mammalian-like unicellular apoptosis », European Journal of Cell Biology, vol. 80, no 6, , p. 428–441 (ISSN 0171-9335, DOI 10.1078/0171-9335-00172, lire en ligne, consulté le )
- (en) Brock, Debra A., Tracy E. Douglas, David C. Queller, et Joan E. Strassmann, « Primitive agriculture in a social amoeba. », Nature, vol. 469 (7330), , p. 393-396. (PMID 21248849)
- Bloomfield, G. et Pears, C. 2003. Superoxide signalling required for multicellular development of Dictyostelium. Journal of Cell Science, 116(16): 3387–3397.
Références taxinomiques
- (en) Référence BioLib : Dictyostelium discoideum Raper 1935 (consulté le )
- (en) Référence Catalogue of Life : Dictyostelium discoideum Raper, 1935 (consulté le )
- (en) Référence Index Fungorum : Dictyostelium discoideum Raper 1935 (+ MycoBank) (consulté le )
- (en) Référence MycoBank : Dictyostelium discoideum Raper (consulté le )
- (en) Référence NCBI : Dictyostelium discoideum (taxons inclus) (consulté le )
- (en) Référence uBio : Dictyostelium discoideum Raper 1935 (consulté le )
Voir aussi
Bibliographie
- Jason S. Kinga et Robert H. Insalla, « Chemotaxis: finding the way forward with Dictyostelium », Trends in Cell Biology, vol. 19 (10), pp. 523-530 (2009).
- Gad Shaulsky et Richard H. Kessin, « The cold war of the social amoebae », Current Biology, vol. 17 (16), pp. R684-R692 (2007).
- Bloomfield, G. et Pears, C. 2003. Superoxide signalling required for multicellular development of Dictyostelium. Journal of Cell Science, 116(16): 3387–3397.
- Tatischeff, I., Petit, P.X., Grodet, A., Tissier J.P., Duband-Goulet, I. et Ameisen, J.C. 2001. Inhibition of multicellular development switches cell death of Dictyostelium discoideum towards mammalian-like unicellular apoptosis. European Journal of Cell Biology.
Liens externes
- http://cgdc3.igmors.u-psud.fr/microbiologie/partie1/chap3_30_myxomycota.htm
- http://microbewiki.kenyon.edu/index.php/Dictiostelida
- http://www.sanger.ac.uk/Projects/D_discoideum/
- dictybase.org/
 Portail de la microbiologie
Portail de la microbiologie  Portail de la mycologie
Portail de la mycologie