Différenciation de niche
La différenciation de niche (synonymes : ségrégation de niches, séparation de niches et partition de niches) est une locution, appliquée au domaine de l'écologie, qui se réfère au processus par lequel une sélection naturelle conduit des espèces concurrentes à utiliser des ressources ou des niches différentes. Ce processus permet à deux espèces de partager certaines ressources sans qu'une espèce n'ait à rivaliser avec l'autre comme dicté par le principe d'exclusion compétitive; ainsi, la coexistence est obtenue par la différenciation des niches écologiques réalisées. La séparation des niches ne peut se produire que s'il y a suffisamment d'espace géographique et écologique pour les organismes puissent s'y développer[3].
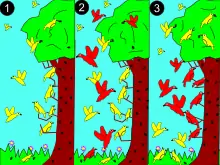
2: une espèce plus grande (rouge) entre en compétition pour les ressources alimentaires.
3: la rouge plus efficace dans l'utilisation des ressources des strates moyennes, exclut la jaune qui est refoulée dans deux nouvelles niches, la strate supérieure et inférieure des arbres
Le principe d'exclusion est bien mis en évidence chez cinq espèces de fauvettes consommant des insectes dans une partie différente[1] des épicéas : elles évitent la compétition par une ségrégation spatiale de leurs niches[2].
La différenciation de niche est un processus qui se produit de plusieurs manières et à plusieurs échelles temporelles et spatiales. Dans la plupart des cas, la différenciation de niche a créé une relation entre deux espèces où la concurrence actuelle est faible ou inexistante. Pour cette raison, la présence de la différenciation de niche peut être méthodologiquement difficile à prouver ou à réfuter. L'absence de preuves de la concurrence actuelle ou passée peut brouiller la distinction entre 1) deux espèces concurrentielles différenciant leurs niches pour pouvoir coexister plutôt que 2) deux espèces non-concurrentes qui occupent des niches écologiques similaires. Il est important de garder à l'esprit que la différenciation de niche et la concurrence inter-spécifique ne peuvent pas toujours être considérées comme liées.
Un exemple de partage des ressources est celui des sept espèces de lézards anolis vivant dans les forêts humides tropicales qui partagent une alimentation commune, principalement des insectes. Ils évitent la concurrence en occupant différentes sections de la forêt tropicale. Certains vivent sur le sol de feuilles mortes tandis que d'autres vivent dans l'épaisseur des branchages, évitant ainsi une concurrence sur la nourriture dans les différents étages de la forêt. Toutes les ressources peuvent faire l'objet de cloisonnement, par exemple, l'espace, la nourriture, les sites de nidification. Cela réduit la concurrence entre les espèces similaires.
Différenciation de niche : détection et quantification.
L'équation de Lotka-Volterra montre que deux espèces concurrentes peuvent coexister lorsque la concurrence intra-spécifique (au sein de l'espèce) est supérieur à la concurrence inter-spécifique (entre les espèces) (Armstrong et McGehee 1980). Depuis différenciation de niche se concentre la concurrence au sein des espèces, en raison d'une diminution de la concurrence entre espèces, le modèle de Lotka-Volterra prédit que la différenciation de niche de n'importe quel degré se traduira par la coexistence.
En réalité, ceci laisse encore en suspens la question de l'importance de la différenciation nécessaire pour qu'une coexistence puisse exister (Hutchinson 1959). Une vague réponse à cette question est que au plus deux espèces sont similaires, au plus elles doivent avoir une adaptation finement équilibrée à leur environnement pour leur permettre de coexister. Il y a des limites à l'importance de la différenciation des niches nécessaires à une coexistence, et celle-ci peut varier selon le type de ressources, la nature de l'environnement et le l'importance de la variation aussi bien à l'intérieur qu'entre les espèces.
Pour répondre aux questions sur la différenciation de niche, il est nécessaire pour les écologues d'être en mesure de détecter, mesurer et quantifier les différents créneaux de coexistence et compétition des espèces. Cela se fait souvent par une combinaison d'études détaillées d'histoire naturelle, des expériences contrôlées (pour déterminer la force de la concurrence), et des modèles mathématiques (1982 Strong, Leibold 1995). Pour comprendre les mécanismes de différenciation de niche et de concurrence, il faut recueillir beaucoup de données sur la façon dont les deux espèces interagissent, comment elles utilisent leurs ressources, et le type d'écosystème dans lequel elles vivent entre autres. En outre, plusieurs modèles mathématiques existent pour quantifier l'ampleur de niche, la concurrence et la coexistence (Bastolla et al. 2005). Toutefois, indépendamment des méthodes utilisées, les niches et la concurrence peuvent être vraiment difficiles à mesurer quantitativement, ce qui rend la détection et la démonstration de différenciation de niche difficile et complexe.
Développement de la différenciation de niche
Au fil du temps, deux espèces concurrentes peuvent coexister soit, grâce à la différenciation de niche ou un autre moyen, soit jusqu'à ce que la concurrence disparaisse une espèce s'éteignant localement. Il existe plusieurs théories sur la façon dont la différenciation de niche évolue compte tenu de ces deux résultats possibles.
Référence
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Niche differentiation » (voir la liste des auteurs).
- Une première près de la pointe des branches, une deuxième dans le feuillage dense, une troisième sur les branches inférieures, une quatrième à la partie supérieure des arbres et la cinquième à leur pointe.
- (en) Robert MacArthur, « Population Ecology of Some Warblers of Northeastern Coniferous Forests », Ecology, vol. 39, no 4, , p. 599–619 (DOI 10.2307/1931600).
- (en) S. Sahney, M.J. Benton et P.A. Ferry, « Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land », Biology Letters, vol. 6, no 4, , p. 544–7 (PMID 20106856, PMCID 2936204, DOI 10.1098/rsbl.2009.1024, lire en ligne [PDF])
Voir aussi
Bibliographie
- (en) R.A. Armstrong et R. McGehee, « Competitive exclusion », American Naturalist, vol. 115, , p. 151–170 (DOI 10.1086/283553)
- (en) U. Bastolla, M. Lässig, S.C. Manrubia et al., « Biodiversity in model ecosystems, I: coexistence conditions for competing species », J. Theor. Biol., vol. 235, no 4, , p. 521–30 (PMID 15935170, DOI 10.1016/j.jtbi.2005.02.005, lire en ligne)
- (en) G.E. Hutchinson, « Homage to Santa Rosalia or Why are there so many kinds of animals? », American Naturalist, vol. 93, , p. 145–159 (DOI 10.1086/282070)
- (en) N. Kronfeld-Schor et T. Dayan, « The dietary basis for temporal partitioning: food habits of coexisting Acomys species », Oecologia, vol. 121, , p. 123–8 (DOI 10.1007/s004420050913)
- (en) S.P. Lawler et P.J. Morin, « Temporal overlap, competition, and priority effects in larval anurans », Ecology, vol. 74, no 1, , p. 174–182 (DOI 10.2307/1939512, lire en ligne)
- (en) M.A. Leibold, « The niche concept revisited: mechanistic models and community context », Ecology, vol. 76, no 5, , p. 1371–82 (DOI 10.2307/1938141, lire en ligne)
- (en) G.H. Pyke, « Local geographic distributions of bumblebees near Crested Butte, Colorado: competition and community structure », Ecology, vol. 63, no 2, , p. 555–573 (DOI 10.2307/1938970, lire en ligne)
- (en) A. Shmida et S. Ellner, « Coexistence of plant species with similar niches », Vegetatio, vol. 58, , p. 29–55
- (en) D.R.J. Strong, « Harmonious coexistence of hispine beetles on Heliconia in experimental and natural communities », Ecology, vol. 63, no 4, , p. 1039–49 (DOI 10.2307/1937243, lire en ligne)
- (en) D. Tilman, « Mechanisms of plant competition for nutrients: the elements of a predictive theory of competition », dans J.B. Grace et D. Tilman, Perspectives on Plant Competition, New York, Academic Press, , p. 117–141
Articles connexes
- Conséquences évolutives de la compétition
- Rôle de la phylogénie dans la détermination de la niche écologique
 Portail de la biologie
Portail de la biologie