Double haploïdie
La double haploïdie, aussi appelée haplodiploïdisation, dihaploïdie ou technique des haploïdes doublés, est une technique de sélection variétale consistant à prélever des cellules haploïdes issues de gamètes d'une plante pour provoquer le doublement de leur stock chromosomique afin d'obtenir une lignée stable en seulement deux générations alors que la fixation (souvent incomplète) des principaux caractères sélectionnés par voie traditionnelle nécessite 6 à 8 générations d'autofécondations amenant à plus de 10 ans le temps nécessaire à l’obtention d'une variété.
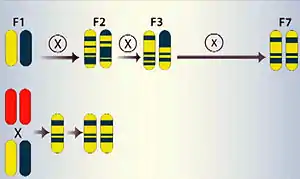
En bas, l'haploïdie générée par un inducteur permet d'obtenir un hybride haploïde doublé parfaitement homozygote dès la seconde génération.
Si on utilise des gamètes mâles (pollen), on parle d'androgenèse. Si on utilise des gamètes femelles (ovules), on parle de gynogenèse.
Une étape de culture in vitro est le plus souvent nécessaire pour régénérer une plante entière car le taux de mutation naturelle de l'haploïdie (stérile) vers la diploïdie (féconde) n'est que de 1 à 10 % selon les variétés.
Contexte
Au cours de la reproduction sexuée, chez les Angiospermes, il se produit une double fécondation. Le grain de pollen contient deux noyaux ou cellules gamétiques mâles qui sont haploïdes : l'une féconde l'oosphère pour former le zygote diploïde qui se développera en un embryon, l'autre fusionne avec le noyau polaire du sac embryonnaire, habituellement déjà diploïde, pour former une cellule triploïde à partir de laquelle se développeront l'albumen et les tissus de réserve.
Exceptionnellement, des anomalies de fécondation peuvent conduire au développement d'un sporophyte à partir d'un seul gamète. Les cellules de cette plante seront alors à n chromosomes (au lieu de 2n) et la plante sera dite haploïde. C'est à partir de cet évènement rare, dont on peut parfois maîtriser la fréquence, que se sont développées de nombreuses études pour l'utilisation de cette anomalie de la reproduction en amélioration des plantes.
Chez le maïs, la présence spontanée de plantes haploïdes au sein d'une population est connue depuis longtemps (Randolph et Stadler, 1929). Leur faible fréquence d'apparition dans différentes descendances, qui varie de 0,5 à 2‰ (Randolph, 1938; Chase, 1949), exclut l’utilisation du système en sélection.
Les premiers travaux sur cette technique ont été initiés par Chase (1949) qui travaillait sur la gynogenèse spontanée et Coe (1959) sur la gynogenèse induite in situ, Kermicle (1969) sur l'androgenèse induite in situ, Miao et al. (1978) sur l'androgenèse in vitro. De nombreuses investigations sur les différentes techniques ont été réalisées depuis et des progrès importants ont été obtenus[1].
Génération de plants haploïdes
On peut obtenir des haploïdes après croisements entre espèces ou entre genres. Il y a fécondation, mais les chromosomes incompatibles du parent pollinisateur sont rejetés naturellement. On peut également provoquer une fécondation anormale à l'aide de pollen dénaturé ou en utilisant des variétés nommées "inducteurs". En 1988, Lashermes et Beckert sélectionnent la lignée de maïs WS14 qui permet l'induction in situ de 2 à 5 % d'individus haploïdes gynogenétiques. Par la suite, d'autres lignées ont été signalées pour leur forte aptitude à induire des plantes haploïdes d'origine gynogénétique[2]. Les variétés de maïs UH400 et Kems atteignent ensuite un taux d'induction de 8 %. En 2010, Procera[3] produit des inducteurs baptisés PHI donnant jusqu'à 15 % d'haploïdes (PHI-3).
Dans tous les cas, on observe le développement d'un embryon haploïde.
Identification des plantes haploïdes
Les plantes haploïdes sont souvent plus petites que leurs géniteurs mais cette miniaturisation n'est pas suffisante pour les sélectionner de façon certaine.
Les gènes dominants de coloration du grain ont, jusqu'à maintenant, été les plus utilisés : lorsque le scutellum de l'embryon n'est pas coloré, il résulte de la division du gamète de la plante induite qui, elle, ne porte pas le gène de coloration. Il y a donc absence de l'allèle dominant de coloration présent chez l'inducteur. Il n'y a pas eu fusion des noyaux mâle et femelle, cet embryon est à n chromosomes, il est donc haploïde.
Ces pigmentations violettes ou pourpres qui apparaissent au niveau de l'embryon ou de la plante, sont dues pour l'essentiel à l'accumulation d'anthocyane[4]. Au moins 14 locus distincts affectent quantitativement et qualitativement l'élaboration et la distribution des pigments anthocyanés. Dans la voie de biosynthèse des anthocyanes, on a pu distinguer des gènes de structure : C2 ou Whp1, A1, A2, Bz1 et Bz2, qui codent des enzymes et agissent de façon linéaire. Chacune des fonctions est nécessaire pour que les pigments se développent. Si une ou plusieurs des trois premières fonctions faiblissent, par la présence d'un homozygote récessif par exemple, les pigments ne sont pas produits. Si un des deux gènes Bz1 ou Bz2 est homozygote récessif, le pigment est de couleur bronze[5].
Il existe des allèles dominants qui peuvent inhiber la biosynthèse des pigments dans la couche à aleurone : C1-I, C2-Idf et In1-D. Coe et Sarkar (1964) utilisent à titre expérimental un caractère de coloration chez le parent femelle et un allèle inhibiteur chez l'inducteur. Chez les grains issus d'une fécondation normale, ni l'embryon, ni l'albumen ne sont colorés. Chez les grains résultant du développement gynogénétique, seul l'embryon est coloré en violet pourpre.
Nanda et Chase (1966) utilisent le caractère purple embryo marker. Ce marqueur dominant est induit par le complexe C2, A1, A2, Bz1, Bz2 C1(R1-nj ou B1-peru), il se traduit par une coloration violet pourpre des grains. Lorsqu'il est présent dans le génome du mâle inducteur, les grains résultant d'une fécondation normale présentent un embryon et un albumen colorés en violet pourpre. En revanche, chez les grains résultant du développement gynogénétique, seul l'albumen est coloré.
Dans la pratique, il est difficile d'identifier un embryon non coloré car dans le grain, il peut être masqué en partie par la couche à aleurone et la coloration du péricarpe. De plus, l'expression de ces marqueurs est fortement influencée par le génotype du parent femelle utilisé, ou même par le processus général de maturation du grain. Toutefois, bien que de fiabilité partielle, ce système permet d'identifier, dans certains types de matériels génétiques, un certain pourcentage des individus haploïdes putatifs. En revanche, pour une étude de comparaison entre différentes techniques, cette identification partielle est susceptible d'induire des biais liés à la non-identification d'un certain nombre de plantes haploïdes.
Retour à l'état fertile diploïde
Pour utiliser en sélection une plante haploïde régénérée, il faut disposer de plantes fertiles et donc diploïdes. L'état haploïde étant instable, l'individu régénéré est parfois diploïde, on parle de doublement spontané du stock chromosomique. Pour le blé, on peut compter 20 à 25 % d'haploïdes doublés spontanément, 60 à 65 % chez l'orge. 10 % chez le maïs (Chase, 1952; Seaney, 1955; Yudin et Khvatova, 1966; Lashermes, 1987) issu d'androgénèse et 50 % pour le maïs issu de gynogénèse.
Sinon, on provoque artificiellement un doublement des chromosomes, le plus couramment par l'action d'un agent chimique, la colchicine.
Les plantes obtenues sont des diploïdes homozygotes : elles possèdent deux copies identiques de chacun de leurs chromosomes et donc portent des paires de gènes ou allèles identiques, d'où leur grand intérêt.
Intérêt de la technique
Lors du retour à l'état diploïde, chaque chromosome de la cellule haploïde de départ est reproduit à l'identique. La technique des haploïdes doublés permet donc d'obtenir rapidement des lignées dont le patrimoine génétique est stable et homozygote.
Elle est souvent utilisée après un croisement, en prélevant des cellules reproductrices sur les plantes de première génération (hybride F1).
En effet, lorsqu'on croise deux plantes ayant des caractères intéressants et complémentaires, on obtient une première génération composée de plantes toutes identiques appelées hybride F1, dont le patrimoine génétique est hétérogène car il provient pour moitié de chacun des parents utilisés pour le croisement. Cette hétérogénéité génère une disjonction des caractères lors des générations suivantes (F2, F3, etc.).
L'obtention de lignées à partir des plantes F1 peut être réalisée de manière naturelle en effectuant des autofécondations successives sur plusieurs générations (7 habituellement) afin d'obtenir des plantes quasi homozygotes. La production d'haploïdes doublés permet de gagner beaucoup de temps puisqu'on obtient un résultat parfaitement homozygote en une seule génération.
Sélection des lignées
Le matériel ainsi fixé est livré au sélectionneur. Le sélectionneur va alors trier les plantes en fonction des critères agronomiques et technologiques recherchés.
La multiplication de ces plantes se fera par autofécondation : tous les descendants seront des copies identiques de leurs parents.
Références
- Jacques BORDES - Création de lignées haploïdes doublées de maïs par gynogenèse induite in situ : amélioration de la méthode et intégration dans les schémas de sélection. 2006
- Zavalishina et Tyrnov, 1992 ; Shastskaya et al., 1994 ; Sarkar et al., 1994 ; Zabirova et al., 1996 ; Deimling et al., 1997 ; Eder et Chalyk, 2002.
- New inducers of maternal haploids in maize. Rotarenco V., Dicu G., State D. and Fuia S. - 2010
- Strauss, 1959
- Neuffer et al., 1997
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire