Folliculogénèse
Chez les animaux, la folliculogénèse est un processus biologique ovarien (dans le cortex ovarien), continu, long (plusieurs mois), très sélectif (99 % des follicules meurent par atrésie) et cyclique consistant en la maturation progressive du follicule ovarien, une enveloppe de cellules somatique contenant un ovocyte immature (ovocyte I).

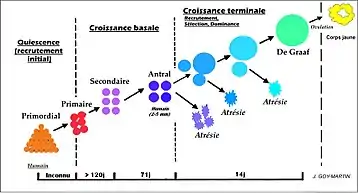
Le début de cette folliculogénèse commence avec le follicule primordial et comprend de nombreuses étapes aboutissant à l'ovulation ou à son involution. Les cellules folliculaires exercent des fonctions à la fois trophiques et endocrines intervenant dans la maturation ovocytaire et l’ovulation.
La maturation du follicule ovarien, au cours de la phase finale de maturation (croissance folliculaire terminale), est stimulée par les gonadotrophines sécrétées par l'antéhypophyse : la FSH (hormone folliculo-stimulante) et l'hormone lutéinisante (LH).
Notons que le nombre de follicules disponibles est fini (contrairement à la spermatogenèse mâle) ce qui entraîne le phénomène de ménopause.
Phase du développement folliculaire humain

La folliculogénèse est un phénomène continu qui débute à la 20e semaine du développement embryonnaire, se poursuit jusqu’à la ménopause (n'est pas interrompu par l’ovulation, ni par la grossesse, ni par d’éventuelles périodes d’anovulation). Elle commence aussi pendant le stade embryonnaire pour d'autres animaux comme la vache. Chez d'autres, comme la souris et la lapine, elle commence après la naissance. Le début de la croissance folliculaire est marquée par le passage de follicule primordiaux.
Elle passe par 5 stades successifs, qui recouvre en fait 2 périodes de signification distincte : une période pré-antrale d'abord, dite de croissance basale et une période antrale ensuite, où le follicule devient sensible au contrôle neuro-endocrinien. Pendant ce temps l’ovocyte primaire est toujours bloqué en prophase l.
Le stade du follicule primordial

La formation du follicule primordial est considérée par beaucoup d'auteurs comme extérieure à la folliculogénèse. Dès que la cellule reproductrice (l'ovocyte) commence sa croissance pendant la vie embryonnaire, elle s'entoure d'une couche unique de cellules (somatiques) aplaties, les cellules folliculaires ou pré-granulosa ou épithéliales, d'origine épithéliale et dérivant des cellules des cordons sexuels. Le follicule primordial mesure alors environ 50 µm, avec un ovocyte de 1er ordre entre 20 et 40µm de diamètre, cet ovocyte est bloqué en 1re phase de division méiotique.
Un stock de réserve de follicules primordiaux est ainsi constitué, et va grandement diminuer tout au long de la vie (300 000 au début de la puberté, 1000 à la ménopause).
À partir de la puberté (chez une femme jeune), 15 à 20 follicules primordiaux entament une phase de croissance par jour vers le stade de follicule primaire, avec un seul (ou deux) arrivant au stade de follicule tertiaire (les autres mourant par atrésie). Le nombre de follicule primordiaux entamant le début de leur croissance chaque jour est proportionnel à l'âge et donc au stock folliculaire.
Phase de quiescence
C'est l'initiation de la croissance folliculaire à proprement parler, cette phase est indépendante des gonadotrophines (LH, FSH).
Le stade du follicule primaire
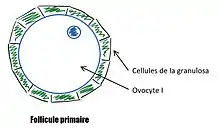
Le passage de follicule primordial à follicule primaire est un processus lent (durée exact mal connu) qui signe le début de la croissance folliculaire. Le follicule et l'ovocyte grossissent, les cellules folliculaires deviennent cubiques et constituent une assise continue autour de l'ovocyte, ce sont les cellules de la granulosa. L'initiation de la croissance est indépendant des gonadotrophines (LH, FSH) mais on retrouve des facteurs impliqués dans le phénomène comme :
- KL, sécrété par les cellules de la granulosa, possède ses récepteurs C-kit au niveau de l’ovocyte. Son absence entraîne l'arrêt de transformation des follicules primordiaux, son augmentation dans le milieu permet d'augmente leur sortie du pool
- LIF et bFGF permettent d'augmenter le nombre de follicules primaires
- AMH (Anti-müllerian hormone) produite par les cellules de la granulosa (pendant le stade primaire et pré-antral) inhibe la croissance des primordiaux en primaires.
Phase de croissance basale
Toute la croissance basale est encore indépendante des gonadotrophines. De plus, la fin de la croissance basale varie entre les espèces, chez la femme elle se termine au stade de follicule antral (diamètre entre 2 et 5 mm). On retrouve des facteurs particuliers impliqués dans cette phase, comme GDF9 (Growth Differentiation Factor 9) et BMP15 (Bone morphogenetic protein15) qui appartiennent à la superfamille des TGFβ (Transforming growth factor beta) et qui sont d'origine ovocytaire (régulent la croissance des follicules primaires et la formation des follicules secondaires). Cette croissance basale dure plus de 191 jours (120 + 71).
Le stade du follicule secondaire

Le follicule primaire continue à se développer pendant plus de 120 jours pour devenir un follicule secondaire. Une couche de molécules glycoprotéiques se forme entre l'ovocyte et les cellules folliculaires, c'est la zone pellucide. La zone pellucide est sécrétée par l'ovocyte (composé de ZP1, ZP2 et ZP3), et intervient dans la reconnaissance spécifique du spermatozoïde. Le stroma du tissu conjonctif commence à former une couche autour du follicule (thèque folliculaire). Le follicule continue d'augmenter de volume et les cellules folliculaires prolifèrent pour former plusieurs couches de cellules. Le follicule fait à ce moment entre 0,06 et 0,15 mm et une lame basale, la membrane de Slavjanski, sépare les cellules de la granulosa du stroma ovarien. Le follicule secondaire mesure environ 200 à 400 µm, avec un ovocyte I de 100 µm de diamètre
Le stade de follicule pré-antral
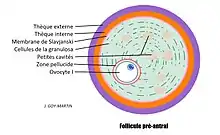
Ce stade est souvent non distingué par de nombreux auteurs et donc assimilé au follicule secondaire. C'est un stade intermédiaire entre le follicule secondaire et le follicule antral. On assiste à la mise en place des ébauches des futures structures du follicule antral (la mise en place complète des structures se fait au stade antral) :
- Les cellules de la granulosa sont plus développées et forment plus de couche,
- La membrane de Slavjanski (membrane basale)
- Les cellules de la future thèque interne
- Les ébauches de la thèque externe (composée de cellules fibreuses)
- Formation progressive de petites cavités (futur antrum)
Le stade du follicule antral
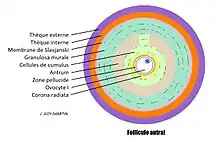
Le passage de secondaire à antral prend environ 71 jours, et se définit par la formation d'une cavité folliculaire : l'antrum. L'antrum se remplit rapidement d'un liquide riche en hormones (œstrogènes) et en AMPcyclique (sécrété par l'ovocyte, les cellules de la granulosa et les thèques). On retrouve en périphérie du follicule antral les mêmes structures histologiques, mais plus évoluées que celles du follicule pré-antral, les thèques se forment complètement et subissent des différenciations fonctionnelles :
- La thèque interne devient très vascularisée (elle sécrétera des androgènes au cours du développement terminal sous l'effet stimulant de la LH)
- La thèque externe devient de nature fibro-musculaire, sa contraction permettra l'éjection de l'ovule hors du follicule lors de l'ovulation.
Pour finir il y a différenciation des cellules de la granulosa en :
- Granulosa murale, au contact de la membrane de Slavjanski,
- Cellules du cumulus oophorus, au contact de la zone pellucide. Une partie de ce cumulus se distingue en corona radiata, directement autour de la zone pellucide. Cette corona radiata (parfois qualifié de cumulus oophorus par certains auteurs) sera expulsée avec l'ovocyte lors de l'ovulation.
Notons que certains auteurs décrivent l’apparition de la corona radiata au stade de follicule de DeGraaf, ou encore entre le stade antral et DeGraaf.
L'ovocyte fait déjà 80-90 % de sa taille finale et le follicule fait, au début de la phase de recrutement, entre 2 et 5 mm.
Développement terminal
Cette dernière phase dépendante des gonadotrophines (LH et FSH) dure 2 semaines (de J0 = début du cycle menstruel, à J14 = ovulation), et commence dès la régression du corps jaune du précédent cycle. Le follicule fait dans les 2 à 5 mm chez la femme jusqu’à l’ovulation. Ce développement (ou croissance) terminal se découpe en trois événements :
- Recrutement
- Sélection
- Dominance
Recrutement
Les follicules antraux gonado-dépendants entrent dans la croissance terminale et forment la cohorte (= ensemble des follicules recrutés). Il y a multiplication très importante des cellules de la granulosa (murale et cumulus) et apparition d'une activité aromatase dans ces cellules. De plus il y a sécrétion d’androgènes par les cellules de thèque interne sous l’effet de la LH, associé à l'apparition de l'activité aromatase, ils sont transformés en œstrogènes par la granulosa.
Notons que l'hormone IGF-1 (ou somatomédine C) permet d'augmenter la taille de la cohorte et potentialise l’action de la LH et de la FSH ; tandis que IGFBP-2 est augmentée dans les cellules de granulosa et de la thèque des follicules en involution[1].
Sélection
La phase de sélection coïncide avec l’apparition des récepteurs à la LH (en plus des récepteurs à la FSH déjà présent) sur les cellules de la granulosa, entraînant une forte production d’inhibine. C'est le début de l'atrésie de quelques follicules.
Dominance
Pendant cette phase de dominance, le follicule dominant (ayant le seuil de réponse à la FSH le plus bas) va évoluer vers le stade préovulatoire (follicule de De Graaf). Pendant la dominance il y a croissance et maturation du follicule dominant, et régression par atrésie des autres follicules de la cohorte (et blocage du recrutement de nouveaux follicules).
Le stade du follicule de De Graaf
Le follicule passant la phase de dominance devient un follicule de De Graaf (ou follicule tertiaire ou follicule mûr ou follicule pré-ovulatoire). Le follicule de De Graaf est un follicule antral plus mature, plus gros, avec des récepteur à la LH et FSH (théorie bicellulaire) et prêt pour l'ovulation.
Au sein du follicule, l'ovocyte I devient l'ovocyte II sous l'effet de la décharge ovulante de LH, et reste bloqué en métaphase II (de la deuxième division méiotique) dans l'attente d'une éventuelle fécondation (voir ovogenèse). L'ovulation survient 35 à 36 heures après la décharge ovulante de LH.
À la suite du pic de LH on assiste à :
- Un arrêt des divisions cellulaires des cellules de la granulosa,
- Un étirement et aplatissement des cellules de l’épithélium ovarien,
- Une contraction du follicule grâce à la thèque externe,
- Une rupture folliculaire,
- Une dissociation des cellules du cumulus oophorus (avec sécrétion importante d’acide hyaluronique sous le contrôle de l’ovocyte),
- Une dissociation des cellules sous-jacentes, de la granulosa, des thèques et de l’albuginée,
- Une fuite du liquide folliculaire,
- La formation du corps jaune.
Le corps jaune
Le corps jaune fait partie de la phase lutéale du cycle menstruel et est issu de la transformation des cellules du follicule de De Graaf après l’ovulation. Sa durée de vie est déterminée (à partir de j14) et varie s'il y a fécondation ou non.
Il sécrète de la progestérone et de l’estradiol sous le contrôle de la LH et des peptides comme des activines et l'inhibine A (l’inhibine A fait partie du rétrocontrôle de la FSH et inhibe ainsi la croissance des autres follicules).
Réserve folliculaire/ovarienne
La réserve folliculaire (liée avec la réserve ovarienne) est composée à plus de 95 % par des follicules primordiaux. Il y a environ 7 millions de follicules dans les ovaires dès la 20e semaine du développement embryonnaire, puis seuls 1 à 2 millions seront encore présents à la naissance, et le stock disponible à la puberté ne sera plus que d’environ 300 000 follicules.
À partir de quarante ans il y a accélération de la dégénérescence des follicules primordiaux (atrésie folliculaire). Lorsque ce stock de follicules est épuisé (vers 1000 follicule restant), on parle de ménopause. Cette atrésie folliculaire est régulée par le facteur pro-apoptotique Bax et le facteur anti-apoptotique Bcl-2. L'inactivation de Bax entraîne la multiplication par 3 du nombre de follicules primordiaux, tandis que l'inactivation de Bcl2 entraîne leur diminution.
Pour calculer la réserve ovarienne, une mesure du taux de AMH est réalisée. L'AMH est corrélée au nombre de follicules antraux et est donc un bon marqueur du pool folliculaire.
Mais cette description n'est pas tout à fait juste, puisque que des chercheurs ont mis au jour des cellules souches ovariennes d'abord chez la souris en 2004, puis chez la femme en 2012, ouvrant ici le champ à des traitements contre l’infertilité pathologique et physiologique féminine.
En considérant une sortie du pool de 20 follicule primordiaux par jour, une estimation facilement réalisable indique en moyenne 41 années de production ovocytaire (300 000 / 20*365), et donc une ménopause survenant vers 54 ans (41 + 13[2]). En réalité, l'âge moyen de la ménopause en France est de 51 ans[3], suggérant une sortie du pool journalière moyenne de 21,6 follicules primordiaux.
Notes et références
- D. Monniaux, B. Mandon-Pépin et P. Monget, « L'atrésie folliculaire, un gaspillage programmé. », M/S. Médecine sciences, vol. 15, no 2, , p. 157-66 (ISSN 1958-5381, DOI 10.4267/10608/1306, lire en ligne, consulté le )
- « L’âge aux premières règles - Focus - Les mémos de la démo - Ined - Institut national d’études démographiques », sur www.ined.fr (consulté le ).
- « Ménopause : causes, symptômes, âge et solutions », Santé Magazine (consulté le ).
Voir aussi
Liens externes
- Article pour comprendre en détail la croissance folliculaire
- http://www.natisens.com/fonction-de-ovaire-la-folliculogenese/
- http://www.santepublique-editions.fr/objects/l-ovogenese.pdf
- https://www.ebiologie.fr/cours/s/22/la-reproduction-sexuee-ovogenese-et-folliculogenese
- http://www.aly-abbara.com/livre_gyn_obs/images/gif/follicules-ovogenese.html
- http://www.medecine.ups-tlse.fr/desc/fichiers/Folliculogenese%20et%20regulations%20intra-ovariennes.pdf
- http://www.lafed-um1.fr/statique/upload/tuteur/Fiche_de_ry_vision_BDR_3590d42b4d.pdf
 Portail de la médecine
Portail de la médecine  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire  Portail de la physiologie
Portail de la physiologie