Hétérocyste
L'hétérocyste est une cellule fixatrice d’azote, transparente, à paroi épaisse, qui se forme dans les filaments de certaines cyanobactéries comme Nostoc punctiforme, Cylindrospermum stagnale ou Anabaena sphaerica. Ces cellules fixent l'azote à partir du diazote (N2) de l'air en utilisant l'enzyme nitrogénase. Cela permet de fournir aux cellules l'azote nécessaire pour leurs biosynthèses[1]. La nitrogénase est inactivée par l'oxygène. Les hétérocystes doivent mettre en place un environnement anaérobie ou microaérobie. Ainsi, la structure et la physiologie particulière des hétérocystes nécessite un changement global de l'expression génétique. Par exemple, les hétérocystes :
- Produisent une paroi plus épaisse, notamment avec des glycolipides qui forment une barrière hydrophobe à l'oxygène
- Produisent la nitrogénase et les autres protéines impliquées dans la fixation de l'azote
- Dégradent le photosystème II, qui produit de l'oxygène
- Augmentent l'expression d'enzyme de la glycolyse
- Produisent des protéines qui dégradent l'oxygène
- Contiennent des composés comme les cyanophycines qui réduisent la diffusion entre les cellules
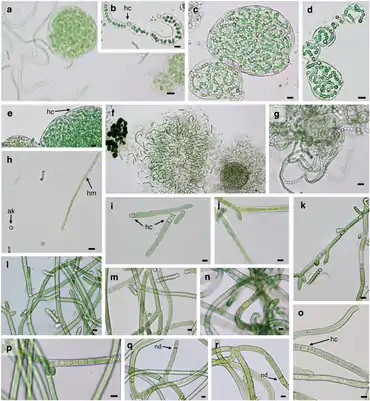
A–F: Nostoc commune G–H: Nostoc calcicola
I–M: Tolypothrix distorta N–R: Scytonema hyalinum
Scale bar = 10 µm , hc, heterocyst, ak, akinete, hm, hormogonium, nd, necridia
Les cyanobactéries peuvent souvent fixer le carbone sous forme de sucres lors de la photosynthèse. L'absence de photosystème II, qui clive l'eau, empêche la réalisation de la photosynthèse au sein des hétérocystes. Ainsi, les cellules végétatives doivent fournir des sucres aux hétérocystes, sous la forme de saccharose. Le carbone et l'azote fixés sont échangés par des canaux présents entre les cellules du filament. Les hétérocystes maintiennent le photosystème I, ce qui leur permet de produire de l'ATP par phosphorylation cyclique.
On trouve un hétérocyste toutes les 9 à 15 cellules, ce qui constitue un motif retrouvé le long d'un filament. L'intervalle entre deux hétérocystes reste approximativement constant malgré la division cellulaire. Les filaments bactériens peuvent être considérés comme des organismes pluricellulaires avec deux types cellulaires distincts et interdépendants. De tels comportements sont assez rares chez les procaryotes mais pourraient correspondre à un des premiers exemples d'apparition de multicellularité au cours de l'évolution. La différenciation en hétérocyste est un phénomène irréversible. En outre, certaines bactéries présentant des hétérocystes peuvent se différencier en cellules ressemblant à des spores appelées akinètes ou en cellules motiles appelées hormogonie, ce qui fait des cyanobactéries des procaryotes avec des phénotypes très variés.
Étymologie
Le terme hétérocyste vient du grec ancien : ἕτερος, (héteros, « autre ») et de cystis, « sac ».
Expression génétique
Dans des environnements pauvres en azote, la différenciation en hétérocyste est induite par le régulateur transcriptionnel NctA. NctA influence la différenciation des hétérocystes par le biais de protéines de signalisation impliquées dans le processus de différenciation. Par exemple, NctA contrôle l'expression de plusieurs gènes dont HetR, qui est un gène essentiel pour la différenciation en hétérocyste[2]. Cette protéine est essentielle vu qu'elle permet d'augmenter la transcription d'autres gènes comme hetR, patS ou hepA en se fixant sur leurs promoteurs et agissant alors comme facteur de transcription. Il est également important de noter que l'expression de nctA et de herR sont dépendantes l'une de l'autre et leur présence va promouvoir la différenciation en hétérocyste. Il a également été récemment montré que d'autres gènes tels que patA ou hetP régulent la différenciation en hétérocystes[3]. PatA permet de déterminer l'intervalle entre les hétérocystes au sein du filament et joue un rôle dans la division cellulaire. PatS influence le motif des hétérocystes en inhibant la différenciation en hétérocyste quand un groupe de cellules en différenciation s'assemble pour former un pro-hétérocyste (hétérocyste immature)[4]. Le maintien des hétérocystes dépend d'une enzyme appelée HetN. La formation d'hétérocystes est inhibée par la présence de source d'azote fixée, telle que l'ammonium ou le nitrate[5].
Relations symbiotiques
La bactérie peut établir une relation symbiotique avec certaines plantes. Dans de telles relations, la bactérie ne répond pas à la disponibilité en azote, mais aux signaux produits par la plante et stimulant la différenciation en hétérocystes. Jusqu'à 60 % des cellules peuvent alors se différencier en hétérocystes, ce qui permet de fournir de l'azote fixé à la plante qui en échange donne à la bactérie du carbone fixé[5]. Le signal produit par la plante et l'état de différenciation des hétérocystes affectés sont encore inconnus. Il est proposé que le signal symbiotique généré par la plante agisse en amont de l'activation de NctA vu que HetR est requis pour la différenciation. Pour l'association symbiotique avec la plante, on sait que nctA est nécessaire vu qu'une bactérie mutée dans le gène nctA ne peut infecter la plante[6].
Notes et références
- (en) Wolk, C.P., Ernst, A. et Elhai, J., « Heterocyst metabolism and development. », The Molecular Biology of Cyanobacteria, , p. 769-823 (ISBN 978-0-7923-3273-2, DOI 10.1007/978-94-011-0227-8_27)
- (en) Herrero, Antonia, Muro-Pastor, Alicia M. et Flores, Enrique, « Nitrogen Control in Cyanobacteria », Journal of Bacteriology, , p. 411-425 (ISSN 0021-9193, PMCID 94895, DOI 10.1128/JB.183.2.411-425.2001)
- (en) Higa, Kelly C. et Callahan, Sean M., « Ectopic expressio of hetP can partially bypass the need for hetR in heterocyst differentiation by Anabaena sp. strain PCC 7120 », Molecular Microbiology, , p. 562-574 (ISSN 1365-2958, PMID 20545862, DOI 10.1111/j.1365-2958.2010.07257.x)
- (en) Orozco, Christine C., Risser, Douglas D. et Callahan, Sean M., « Epistasis Analysis of Four Genes from Anabaena sp. Strain PCC 7210 Suggests a Connection between PatA and PatS in Heterocyst Pattern Formation », Journal of Bacteriology, , p. 1808-1816 (ISSN 0021-9193, PMCID 1426565, DOI 10.1128/JB.188.5.1808-1816.2006)
- lee, Robert Edward. Phycology.
- (en) Meeks, JC et Elhai, J, « Regulation of Cellular Differenciation in Filamentous Cyanobacteria in Free-Living and Plant-Associated Symbiotic Growth States », Microbiology and Molecular Biology Reviews, , p. 94-121 (PMID 11875129, PMCID 120779, DOI 10.1128/MMBR.66.1.94-121.2002)