Histone acétyltransférase
L'histone acétyltransférase, abrégée en HAT, est une acétyltransférase qui catalyse la réaction :
- acétyl-CoA + histone CoA + acétylhistone.

Pour les articles homonymes, voir HAT.
| N° EC | EC |
|---|---|
| N° CAS |
| IUBMB | Entrée IUBMB |
|---|---|
| IntEnz | Vue IntEnz |
| BRENDA | Entrée BRENDA |
| KEGG | Entrée KEGG |
| MetaCyc | Voie métabolique |
| PRIAM | Profil |
| PDB | RCSB PDB PDBe PDBj PDBsum |
| GO | AmiGO / EGO |
Cette enzyme est associée à l'activation de la transcription, notamment en agissant sur le remodelage et la décondensation de la chromatine, permettant ainsi l'exposition de nombreux sites de liaisons pour l'ARN polymérase II et pour les protéines de régulation de la transcription.
Contexte : l'organisation du génome eucaryote
Le matériel génétique chez les eucaryotes est situé dans le noyau où une forte compaction est nécessaire afin de contenir tout l’ADN dans le noyau (6µm de diamètre). À titre d’exemple, l’ADN est long de 2 mètres une fois « déroulé » et atteint une compaction de 10 000 fois lors de la formation du chromosome métaphasique. L'ADN se présente sous deux formes principales : l’euchromatine et l’hétérochromatine. L’euchromatine est la moins répandue (30 % du génome), et correspond aux séquences d’ADN exprimées par la cellule alors que la forme hétérochromatine (70 % du génome) est très condensée et porte les séquences non exprimées par la cellule. L’ADN s’enroule autour de 4 protéines histones présentes en 2 copies chacune (code histone) constituant un nucléosome. Du niveau de compaction de ces nucléosomes dépend l’état d’activation des gènes. En effet, plus les nucléosomes sont condensés, moins la transcription est facilitée. En d’autres termes, la forte condensation de l’ADN empêche la fixation de facteurs activateurs de la transcription ainsi que le recrutement de l’ARN polymérase permettant la transcription des gènes. De nombreuses enzymes sont impliquées dans la régulation de l’état de condensation de la chromatine. C’est le cas de l’histone acétyltransférase (HAT) qui intervient dans la décondensation de l'ADN.
Rôles de l'histone acétyltransférase
L'histone acétyltransférase (HAT) est une enzyme intervenant dans le remodelage de la chromatine. Son action consiste à acétyler les résidus lysine présents sur les queues N-terminales des histones. Cette acétylation va alors permettre à la chromatine de se décompacter et d'exposer des sites de liaison pour l'ARN polymérase II et pour les protéines de régulation de la transcription (facteurs de transcription). L'acétylation des extrémités N-terminales des histones par des enzymes possédant une activité HAT constitue également une marque épigénétique essentielle dans le concept de code des histones. En effet, la lysine acétylée devient alors le site de reconnaissance spécifique de protéines à bromodomaines (en), point de départ du recrutement d'une multitude de facteurs régulant la transcription.
Mode d'action
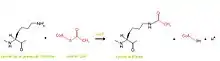
Les queues N-terminales des protéines histones liées à l'ADN et formant le nucléosome peuvent subir des modifications post-traductionelles. L'histone acétyltransférase (HAT) va, comme son nom l’indique, transférer un groupement acétyle sur certains résidus lysines des queues N-terminales. L'enzyme HAT catalyse la création d'un groupe amide non chargé par l'addition d'un groupement acétyle (provenant de l'acétyle Coenzyme A) sur le groupement ammonium de la lysine (figure 1).
La lysine, possédant un groupement ammonium chargé positivement à pH neutre (entraînant une grande affinité entre l'histone et l'ADN) ne sera plus chargée après l'intervention de l’HAT. En effet, le groupement ammonium est remplacé par un groupement amide neutre à la suite de la fixation du groupement acétyle. De ce fait il y a une perte d'affinité entre l'histone et l'ADN due à la perte de la charge positive. Cette perte d'affinité provoque un relâchement du complexe protéines/ADN, dévoilant ainsi de nouveaux segments d'ADN nécessaires au mécanisme de la transcription. De plus le groupement acétylé, aussi appelé acétyllysine est reconnu par les bromodomaines présents sur les protéines des moteurs de remodelages de la chromatine, ces dernières utilisent l’énergie libre d’hydrolyse de l’ATP afin de déplacer les positions des nucléosomes le long de l’ADN et induisent d’autres changements conformationnels dans la chromatine. La partie de l’ADN ainsi dépliée sera alors exposée aux différents facteurs de la transcription ainsi qu’à l’ARN polymérase II nécessaire à la transcription.
Les histones désacétylases (HDAC)
Il existe aussi des enzymes catalysant la réaction inverse, c’est-à-dire permettant la condensation de la chromatine, par désacétylation des groupements acétyllysine, c’est l’histone désacétylase. De ce fait cette enzyme joue un rôle de répresseur cette fois ci dans la régulation de l’activité transcriptionnelle.
Voir aussi
Bibliographie
- Garrett & Grisham, Biochimie, traduction de la 2e édition américaine par Bernard Lubochinsky, édition DeBoeck Université, 2000
- L. Stryer, J. M. Berg et J. L. Tymoczko, Biochimie, Editions Flammarion médecine-sciences, 5e édition, 2003
- J.H. Weil, Biochimie générale, 11e édition, Editions Dunod
Articles connexes
 Portail de la biochimie
Portail de la biochimie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire