Nestedness
L'imbrication (ou emboîtement ou Nestedness) désigne en écologie un concept apparu dans les années 1990 dans le cadre de la biogéographie pour décrire la manière dont les écosystèmes sont construits, ou plus exactement "structurés" dans l'espace[1]. Il s'agit "patron de répartition" d'espèces où des communautés pauvres en espèces (à faible diversité spécifique) sont un sous-ensemble non-aléatoire de communautés d'espèces plus riches en nombre d'espèces (dans une région donnée).
A titre d'exemple on peut imaginer une série d'îles, classées selon leur distance à un continent homogène. Si le continent abrite toutes les espèces, la première île n'abritera qu'un sous-ensemble des espèces continentales, et l'île n°2 n'abritera qu'une partie (un sous-ensemble) des espèces de l'île n°1 et ainsi de suite. Ce type de système est dit "parfaitement imbriqué".
Cette notion d'imbrication est l'un des outils permettant d'analyser, décrire et mesurer la "structure" d'un système écologique, permettant par exemple de mieux comprendre comment des sous-populations s'organisent géographiquement via des phénomènes de turnover[2], colonisation/extinctions autour d'une population-source et de "puits écologiques", et parfois de "pièges écologiques", au sein d'une métapopulation[3].
Elle est habituellement appliquée aux systèmes espèces-sites (c'est-à-dire s'intéressant à la répartition des espèces entre les sites, comme dans l'exemple précédent îles-continent (cf. théorie de l'insularisation écologique[4]), ou en fonction d'autres facteurs que l'éloignement, par exemple selon l'altitude, la profondeur marine, le degré de trophie...).
Mais l'imbrication peut aussi s'appliquer aux réseaux d'interaction entre espèces (pour décrire des interactions entre espèces telles que les réseaux hôtes-parasites, plantes-pollinisateurs, prédateurs-proies, etc.).
On dit qu'un système (habituellement représenté comme une matrice) est "imbriqué" si les éléments qui contiennent peu d'items (ex : site avec peu d'espèces, espèces avec peu d'interactions) ont un sous-ensemble d'éléments avec plus d'éléments.
Comprendre et connaître le degré d'imbrication d'espèces, de communautés ou d'habitats a des implications importantes pour la conservation de la nature[5].
Mesure et représentation du degré d'imbrication
De nombreuses mesures ont été proposées pour quantifier la nestedness au sein des réseaux d’interactions mutualistes. Parmi ces propositions, deux sont particulièrement utilisées: la température [4] et la NODF (Nestedness metric based on Overlap and Decreasing Fill)[6].

Mesure de la température
Atmar et Patterson en 1993 ont proposé d'utiliser la métaphore de la température[7].
Le système le plus riche en espèce ou en interaction est le plus chaud (et peut être coloré en couleur chaude sur une carte ou une autre forme de représentation du système étudié). Les variations géographiques de cette "pseudo-température" donne une indication sur l'ordre probable dans lequel les extinctions d'espèces se produiront dans le système (ou inversement des indications sur les sens probables de colonisation d'un système par les espèces d'un autre système).
Plus le système est «froid» plus l'ordre d'extinction sera fixe et plausiblement facile à anticiper. Inversement plus le système est "chaud" plus les extinctions y sembleront aléatoires et imprévisibles. Dans cette version de la théorie, les températures peuvent s'échelonner de 0° (système le plus froid et absolument fixe) à 100° (absolument aléatoire).
Dans les années 2000 on développe des algorithmes de calcul de l'imbrication[8], pour parer aux limites de la première "calculatrice de température d'imbrication"[9] qui pour diverses raisons n'était pas mathématiquement satisfaisante (pas de solution unique, pas de conservation)[10],[11]. Un logiciel (BINMATNEST) est disponible auprès des auteurs sur demande et auprès du Journal of Biogeography pour corriger ces déficits[12]. En outre, ANINHADO résout les problèmes de grande taille de la matrice et le traitement d'une grande le nombre de matrices randomisées : il implémente plusieurs modèles nuls pour estimer l'importance de l'imbrication[11].
En 2009 Bastolla et al. ont introduit une mesure simple de l'imbrication basée sur le nombre de voisins communs pour chaque paire de nœuds[13]. Ils soutiennent que cela peut aider à réduire la concurrence effective entre les nœuds dans certaines situations. Par exemple, deux espèces d'insectes peuvent «s'entraider» en pollinisant le même sous-ensemble de plantes, réduisant ainsi la mesure dans laquelle elles se nuiraient entre elles. Les auteurs suggèrent que cet effet est à l'origine d'une corrélation entre le degré d'imbrication et la diversité dans les écosystèmes plantes-pollinisateurs.
Johnson et al. ont montré que cette mesure ne tient pas compte de l'effet désiré[14].
Ces auteurs proposent une version affinée de la mesure, et continuent à étudier comment certaines propriétés de certains "réseaux écologiques" affectent l'imbrication (la qualité des habitats en particulier[15]).
Mesure du NODF
La NODF (pour Nestedness metric based on Overlap and Decreasing Fill) est une métrique proposée par ALMEIDA-NETO et ses collaborateurs en 2008. C'est également l’une des méthode les plus utilisées afin de mesurer la nestedness.
La NODF se mesure tel que[16]:

Avec = le nombre de colonnes

= le nombre de lignes

= la moyenne du degré de nestedness au sein des colonnes

= la moyenne du degré de nestedness au sein des lignes

Les valeurs prises vont de 0 (nestedness nulle) à 100 (parfaite nestedness).
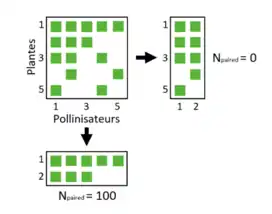
Sous l’hypothèse d’un réseau très imbriqué, la matrice optimisée présente un nombre important d’interactions sur les lignes du haut et les colonnes de gauche (espèces les plus généralistes). Ce nombre diminue avec les espèces de plus en plus spécialistes (colonnes de droite et lignes du bas). Ce phénomène de diminution du nombre d'interactions est appelé DF (Decreasing Fill), c’est la première propriété sur laquelle repose la mesure NODF.
Dans le calcul de Ncol, chacune des colonnes est comparée. Si la propriété n’est pas validée (pas de diminution d’interactions entre les deux colonnes), le degré de nestedness (Npaired) est égal à 0. Dans le cas inverse, Npaired est égal à PO (Paired Overlap), c’est la seconde propriété de la mesure. Elle correspond au nombre d’interactions communes entre deux colonnes, par rapport au nombre total d’interactions sur la colonne de droite (et de même avec les lignes).
Cas particuliers
Cette approche a beaucoup été utilisée pour les îles vraies, mais elle a aussi été déclinée pour
- la flore vivant ou survivant dans le contexte de la fragmentation forestière (naturelle ou artificielle)[17]
- des mares de ponte d'amphibiens (groupe taxinomique en forte régression dans le monde)[18]
- des lacs plus ou moins isolés (en zone boréale par exemple[19])
- des cours d'eau. On a depuis longtemps constaté que les communautés d'espèces s'y structurent notamment en fonction de la largeur/profondeur du segment du cours d'eau considéré (qui dépend aussi souvent de son altitude) [20]3 chercheurs finlandais ont étudié le degré d'emboîtement d'assemblages d'insectes dans des ruisseaux de tête de bassin versant de zone boréale (63–70° N, 23–29° E) en cherchant quels étaient les déterminants des emboitements, en partant de l'hypothèse que l'emboîtement des caractéristiques de l'habitat et de la structure des niches (plus ou mois fractal dans le cas des cours d'eau) crée un emboîtement chez les organismes qui les colonisent. Cette hypothèse a été testée pour 8 bassins versants et a été confirmée dans ces 8 cas par le "calculateur de température d'emboîtement" et dans 5 cas sur la base d'une analyse de divergence.
Dans le bas du bassin le degré d'emboîtement était assez faible, comme le suggérait les valeurs élevées de "température" de la matrice. Dans la matrice d'emboîtement l'ordre des sites était significativement corrélé à la taille des ruisseaux dans 2 des régions et aux gradients environnementaux dans 5 des régions. L'emboîtement semblait surtout régi par la taille des ruisseaux et par les gradients environnementaux locaux. Les liens entre emboîtement et gradients environnementaux évoquent pour certaines zones une structure en niches emboitées liée aux traits correspondant à leurs réponses aux gradients environnementaux. Dans ce type de contexte ces liens semblent donc très spécifiques à la région, évoquant des patrons d'emboîtement des sous-ensembles très dépendants du contexte biogéographique[21].
Références
- Boecklen, W. J., (1997). Nestedness, biogeographic theory, and the design of nature reserves. Oecologia, 112: 123–142.
- Leibold M.A. & G.M Mikkelson (2002). Coherence, species turnover, and boundary clumping: Elements of meta-community structure. Oikos, 97: 237–250.
- Heino J (2005). Metacommunity patterns of highly diverse stream midges: Gradients, chequerboards and nestedness, or is there only randomness? Ecological Entomology, 30: 590–599
- Atmar W. & B.D. Patterson (1993). The measure of order and disorder in the distribution of species in fragmented habitat. Oecologia, 96: 373–382
- Fleishman, E. & R. MacNally, 2002. Topographic determinants of faunal nestedness in Great Basin butterfly assemblages: Applications for conservation planning. Conservation Biology, 16:422–429
- Mário Almeida-Neto, Paulo Guimarães, Paulo R. Guimarães et Rafael D. Loyola, « A consistent metric for nestedness analysis in ecological systems: reconciling concept and measurement », Oikos, vol. 117, no 8, , p. 1227–1239 (ISSN 0030-1299, DOI 10.1111/j.0030-1299.2008.16644.x, lire en ligne, consulté le )
- Patterson and Atmar et Bruce D. Patterson, « The measure of order and disorder in the distribution of species in fragmented habitat », Oecologia, vol. 96, no 3, , p. 373–382 (DOI 10.1007/BF00317508)
- ex : Rodríguez-Gironés, M. A. & L. Santamaria (2006). A new algorithm to calculate the nestedness temperature of presenceabsence matrices. Journal of Biogeography, 33: 924–935.
- Atmar, W. & B. D. Patterson (1995). The Nestedness Temperature Calculator: A Visual Basic Program, Including 294 Presence—Absence Matrices. AICS Research Inc., Chicago, Illinois. [Online] URL: http://aics-research.com/nestedness/tempcalc.html
- M. A. Rodríguez-Gironés et L. Santamaría, « A new algorithm to calculate the nestedness temperature of presence–absence matrices », Journal of Biogeography, vol. 33, no 5, , p. 924–935 (DOI 10.1111/j.1365-2699.2006.01444.x)
- (en) Paulo R. Guimarães Junior et Paulo Guimarães, « Improving the analyses of nestedness for large sets of matrices », Environmental Modelling and Software, vol. 21, no 10, , p. 1512–1513 (ISSN 1364-8152, DOI 10.1016/j.envsoft.2006.04.002)
- https://doi.org/10.1111/j.1365-2699.2006.01444.x
- U. Bastolla, M. A. Fortuna, A. Pascual-García, A. Ferrera, B. Luque et J. Bascompte, « The architecture of mutualistic networks minimizes competition and increases biodiversity », Nature, vol. 458, , p. 1018–1020 (PMID 19396144, DOI 10.1038/nature07950)
- S. Johnson, V. Domínguez-García et M. A. Muñoz, « Factors Determining Nestedness in Complex Networks », PLoS ONE, vol. 8, , e74025 (DOI 10.1371/journal.pone.0074025)
- Hylander, K., C. Nilsson, B. G. Jonsson & T. Göthner, (2005). Differences in habitat quality explain nestedness in a land snail metacommunity. Oikos, 108: 351–361.
- Beckett, Stephen James., Nestedness and modularity in bipartite networks (OCLC 1182009912, lire en ligne)
- Honnay, O., M. Hermy & P. Coppin, (1999) Nested plant communities in deciduous forest fragments: Species relaxation or nested habitat ? Oikos, 84: 119–129
- Hecnar, S. J. & R. T. M'Closkey, (1997). Patterns of nestedness and assemblage composition in a pond-dwelling amphibian fauna. Oikos, 80: 371–381.
- Heino J. & T. Muotka, (2005) Highly nested snail and clam assemblages in boreal lake littorals: Roles of isolation, area and habitat suitability. Écoscience, 12: 141–147
- Brönmark, C., J. Herrmann, B. Malmqvist, C. Otto & P. Sjöström, 1984. Animal community structure as a function of stream size. Hydrobiologia, 112: 73–79
- Jani Heino, Heikki Mykrä & Jaana Rintala (2010) ; Assessing Patterns of Nestedness in Stream Insect Assemblages Along Environmental Gradients ; Ecoscience 17(4):345-355. 2010 https://doi.org/10.2980/17-4-3263
Voir aussi
Bibliographie
- Brualdi, R. A. & J. G. Sanderson, 1999. Nested species subsets, gaps, and discrepancy. Oecologia, 119: 256–264.
- Calme, S. & A. Desrochers, 1999. Nested bird and microhabitat assemblages in a peatland archipelago. Oecologia, 118: 361–370.
- Cook, R. R. & J. F. Quinn, 1995. The influence of colonization in nested species subsets. Oecologia, 102: 413–424
- Fleishman, E. & R. MacNally, 2002. Topographic determinants of faunal nestedness in Great Basin butterfly assemblages: Applications for conservation planning. Conservation Biology, 16:422–429
- Hecnar, S. J. & R. T. M'Closkey, 1997. Patterns of nestedness and assemblage composition in a pond-dwelling amphibian fauna. Oikos, 80: 371–381.
- Heino, J., 2005a. Metacommunity patterns of highly diverse stream midges: Gradients, chequerboards and nestedness, or is there only randomness? Ecological Entomology, 30: 590–599.
- Heino, J., 2005b. Positive relationship between regional distribution and local abundance in stream insects: A consequence of niche breadth or niche position? Ecography, 28: 345–354.
- Heino, J. & T. Muotka, 2005. Highly nested snail and clam assemblages in boreal lake littorals: Roles of isolation, area and habitat suitability. Écoscience, 12: 141–147. BioOne,
- Heino, J., T. Muotka & R. Paavola, 2003. Determinants of macroinvertebrate diversity in headwater streams: Regional and local influences. Journal of Animal Ecology, 72: 425–434.
- Heino, J., H. Mykrä & T. Muotka, 2009. Temporal variability of nestedness and idiosyncratic species in stream insect assemblages. Diversity and Distributions, 15: 198–206.
- Heino, J., T. Muotka, R. Paavola & L. Paasivirta, 2003. Amongtaxon congruence in biodiversity patterns: Can stream insect diversity be predicted using single taxonomic groups? Canadian Journal of Fisheries and Aquatic Sciences, 60: 1039–1049.
- Honnay, O., M. Hermy & P. Coppin, 1999. Nested plant communities in deciduous forest fragments: Species relaxation or nested habitat? Oikos, 84: 119–129.
- Hylander, K., C. Nilsson, B. G. Jonsson & T. Göthner, 2005. Differences in habitat quality explain nestedness in a land snail metacommunity. Oikos, 108: 351–361.
- Jonsson, B. G., 2001. A null model for randomization test of nestedness in species assemblages. Oecologia, 127: 309–313.
- Kadmon, R., 1995. Nested subsets and geographical isolation. Ecology, 76: 458–465.
- Kerr, J. T., A. Sugar & L. Packer, 2000. Indicator taxa, rapid biodiversity assessment, and nestedness in an endangered ecoregion. Conservation Biology, 14: 1726–1734.
- Lomolino, M. V., 1996. Investigating causality of nestedness of insular communities: Selective immigrations or extinctions? Journal of Biogeography, 23: 699–703.
- Malmqvist, B. & Å. Eriksson, 1995. Benthic insects in Swedish lake-outlet streams: Patterns in species richness and assemblage structure. Freshwater Biology, 34: 285–296.
- Malmqvist, B. & P.-O. Hoffsten, 2000. Macroinvertebrate taxonomic richness, community structure and nestedness in Swedish streams. Archiv für Hydrobiologie, 150: 29–54.
- Malmqvist, B. & M. Mäki, 1994. Benthic macroinvertebrate assemblages in north Swedish streams. Ecography, 17: 9–16.
- Malmqvist, B., V. Meisch & A. N. Nilsson, 1997. Distribution patterns of freshwater Ostracoda (Crustacea) in the Canary Islands with regards to habitat use and biogeography. Hydrobiologia, 347: 159–170.
- Malmqvist, B., Y. Zhang & P. H. Adler, 1999. Diversity, distribution and larval habitats of North Swedish blackflies (Diptera: Simuliidae). Freshwater Biology, 42: 301–314.
- Maron, M., R. MacNally, D. M. Watson & A. Lill, 2004. Can the biotic nestedness matrix be used predictively? Oikos, 106: 433–444.
- McAbendroth, J., A. Foggo, S. D. Rundle & D. T. Bilton, 2005. Unravelling nestedness and spatial pattern in pond assemblages. Journal of Animal Ecology, 74: 41–49.
- McCune, B. & M. J. Mefford, 1999. PC-Ord: Multivariate Analysis of Ecological Data. Version 4.25. MjM Software, Gleneden Beach, Oregon.
- McDonald, K. A. & J. H. Brown, 1992. Using montane mammals to model extinctions due to global change. Conservation Biology, 6: 409–415.
- Monaghan, M. T., C. T. Robinson, P. Spaak & J. V. Ward, 2005. Macroinvertebrate diversity in fragmented alpine streams: Implications for freshwater conservation. Aquatic Sciences, 67: 454–464.
- Mykrä, H., J. Heino & T. Muotka, 2007. Scale-related patterns in the spatial and environmental components of stream macroinvertebrate assemblage variation. Global Ecology and Biogeography, 16: 149–159.
- Mykrä, H., T. Ruokonen & T. Muotka, 2006. The effect of sample duration on the efficiency of kick-sampling in two streams with contrasting substratum heterogeneity. Verhandlungen der internationalen Vereinigung für theoretische und angewandte Limnologie, 29: 1351–1355.
- Nilsson, A. N., 1996. Aquatic Insects of North Europe: A Taxonomic Handbook. Volume 1. Apollo Books, Stenstrup.
- Patterson, B. D., 1987. The principle of nested subsets and its implications for biological conservation. Conservation Biology, 1: 323–334.
- Patterson, B. D. & W. Atmar, 1986. Nested subsets and the structure of insular mammal faunas and archipelagos. Biological Journal of the Linnean Society, 28: 65–82.
- Patterson, B. D. & W. Atmar, 2000. Analyzing assemblage composition in fragments. Pages 9–24 in G. Rheinwald (ed.). Isolated Vertebrate Communities in the Tropics. Bonner Zoologische Monographen 46, Bonn.
- Rosenzweig, M. J., 1995. Species Diversity in Space and Time. Oxford University Press, Oxford.
- Sanderson R. A., M. D. Eyre & S. P. Rushton, 2005. The influence of stream invertebrate composition at neighbouring sites on local assemblage composition. Freshwater Biology, 50: 221–231.
- Sokal, R. R. & F. J. Rohlf, 1997. Biometry. 3rd Edition. W. H. Freeman and Company, New York, New York.
- Taylor, C. M. & M. L. Warren, 2001. Dynamics of assemblage composition of stream fish assemblages: Environmental variability and nested subsets. Ecology, 82: 2320–2330.
- Townsend, C. R., A. G. Hildrew & J. Francis, 1983. Community structure in some southern English streams: The influence of physicochemical factors. Freshwater Biology, 13: 521–544
- Ulrich, W. & N. J. Gotelli, 2007. Null model analysis of species nestedness patterns. Ecology, 88: 1824–1831.
- Waite, T. A. & L. G. Campbell, 2006. Controlling the false discovery rate and increasing statistical power in ecological studies. Écoscience, 13: 439–442. BioOne,
- Worthen, W. B., 1996. Community composition and nested-subset analyses: Basic descriptors for community ecology. Oikos, 76: 417–426.
- Wright, D. H., B. D. Patterson, G. M. Mikkelson, A. Cutler & W. Atmar, 1998. A comparative analysis of nested subset patterns of assemblage composition. Oecologia, 113: 1–20
logiciels
- Nestedness Temperature Calculator Program
- « http://www.umk.pl/~ulrichw/Downloads.html#progr9 an Fortran Version correcting some of the initial problems, by Werner Ulrich, Poland »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?)
- ANINHADO, a Nestedness Temperature Calculator Program improving NTC for large matrices and implementing additional null models, by Paulo Guimarães and Paulo R. Guimarães Jr., Brazil
- Weighted Interaction Nestedness Estimator. It works with weighted matrices (i.e. species abundance)
- NeD - Nestedness for Dummies. A user friendly interface for nestedness analysis
 Portail de l’écologie
Portail de l’écologie  Portail de l’environnement
Portail de l’environnement