Écologie des systèmes lotiques
L'écologie de système lotique est l'étude des interactions biotiques et abiotiques dans les flux d'eaux courantes continentales[1]. Avec l'écologie des systèmes lentiques, qui implique des eaux continentales moins dynamiques comme des lacs ou des étangs, ces domaines forment le champ d'études plus générale de l'eau douce ou de l'écologie aquatique.
Les milieux lotiques prennent des formes très diverses, allant du ruisseau peu large au grand fleuve de plusieurs kilomètres de large[2], mais présentent des invariances[3] qui justifient leur étude commune. On observe donc quelques caractéristiques communes rendant l'écologie des eaux coulantes unique par rapport aux autres habitats aquatiques : les flots sont unidirectionnels ; ces systèmes sont dans un état de changement physique continuel ; il y a un haut degré d'hétérogénéité spatiale et temporelle à toutes échelles (microhabitats) ; la variabilité entre les systèmes lotiques est haute ; la « biota » est spécialisée pour vivre dans les conditions des flots.
Facteurs abiotiques importants
Les flux
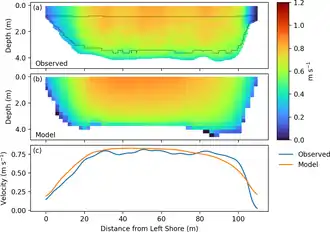
La vitesse intégrée sur les 2 m supérieurs de la colonne d'eau est tracée en c. La ligne pointillée dans le panneau supérieur indique l'étendue des données acquises initialement par un PCED (profileur de courant à effet Doppler) avant extrapolation.
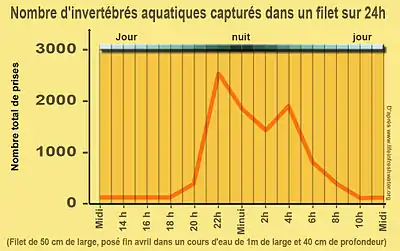
Dans les systèmes lotiques, de nombreux invertébrés se détachent de leur support, ou se laissent volontairement emporter par le courant. Ceci se produit presque toujours dans les périodes crépusculaire et dans les 2 ou 3 heures qui suivent le coucher du soleil et peu avant l’aube. Les poissons et certaines espèces prédatrices semblent également plus actifs à ces périodes. La pollution lumineuse pourrait peut-être perturber ce mécanisme qui semble lié aux variations d'intensité de la lumière perçue par les invertébrés.
Les flux physiques (eau, sédiments en suspension, oxygène dissous...) sont dans les systèmes lotiques l'opérateur abiotique clé (éventuellement influencé par l'homme via ses modes d'aménagement du territoire et d'occupation des sols) qui influencent leur écologie.
La force des flux d'eau peut varier selon le système et ses sous-systèmes. Dans un cours d'eau rapide ou lent, certains remous peuvent presque être vus comme des systèmes lentiques.
La vitesse des flux d'eau peut être variable à l'intérieur des systèmes et entre systèmes différents. C'est typiquement basé sur la variabilité de friction avec le fond et les berges du cours d'eau, la sinuosité, les obstructions (naturelles ou non), et le degré d'inclinaison de la pente[2].
De plus, la quantité d'eau entrant dans le système à partir de précipitations, de la fonte des neiges et/ou d'apports d'eaux souterraines peuvent affecter la vitesse de flux. Les eaux peuvent altérer la forme du lit de la rivière à travers des processus d'érosion, de surcreusement ou au contraire de déposition, créant une variété d'habitats, incluant des courants, glissements et mares[4].
Les flux biologiques (grandes et petites migrations) varient aussi selon la saison et le segment considéré du cours d'eau, de l'amont à l'aval. On observe aussi des variations jour-nuit dans le flux d'individus (invertébrés, poissons) se laissant porter par le courant.
Dans tous les cas, dans la nature, l'échelle du bassin versant est fortement prégnante. Dans un nombre croissant de région du monde, les bassins versants ont été mis en relation par des canaux, ce qui a favorisé de nombreux phénomènes d'invasion biologique. Certaines espèces invasives peuvent aussi être apportées par les cales ou coques de bateaux.
Lumière
La lumière solaire est importante pour les systèmes lotiques. Les UV qu'elle contient contribuent à une « désinfection » naturelle de l'eau. Le soleil est nécessaire à la photosynthèse qui produit l'énergie nécessaire à la productivité primaire. Par l'ombre qu'elle produit, la lumière crée des refuges pour les proies.
La quantité de lumière reçue par un système peut être reliée à une combinaison de variables de flux internes et externes. L'aire entourant un petit ruisseau, par exemple, peut être ombré par des forêts (ripisylves, forêt-galerie, de hautes berges, voire des falaises. De plus grands systèmes de rivière ou fleuve tendent à être large, réduisant l'influence des variables externes. Le soleil gagne de la surface. Certaines espèces jouent également un rôle en freinant l'eau et en ouvrant la rivière au soleil ; ce sont les castors. Les rivières plus turbulentes se chargent en particules. De même les rivières riches en nutriments (phosphates et nitrates en particulier) peuvent abriter une biomasse algale importante. Les particules ou cellules en suspension et le plancton, s'ils sont très présent, atténuent la pénétration de la lumière, d'autant plus que la profondeur augmente[4]. Les facteurs saisonniers et journalier peuvent jouent aussi un rôle dans la distribution lumineuse, avec une modification de l'intensité lumineuse moyenne, mais aussi de l'angle d'incidence (l'angle que forme les rayons lumineux avec l'eau peut mener à une déperdition de lumière à cause de la réflexion. Connue comme la loi de Beer-Lambert, plus l'angle est petit, plus la lumière est réfléchie et la quantité des rayonnements solaires reçus décroit logarithmiquement avec la profondeur[3]). D'autres influences sur la quantité de lumière sont la couverture nuageuse, l'altitude, et la position géographique[5].
Température
La plupart des espèces lotiques sont des poïkilothermes, c'est-à-dire que leur température interne varie avec leur environnement, et c'est pourquoi la température est considérée comme un facteur abiotique clef. L'eau peut être chauffée par radiation en surface et échanger de la chaleur par conduction avec l'air ou le substrat sous-jacent. Les couches d'eau peu profondes sont relativement bien mélangées par agitation de l'eau et conservent donc des températures quasi identiques. L'eau circulant à proximité ou au contact du fond est, quant à elle, beaucoup moins rapide et il se développe souvent une forte différence de température entre la surface et le fond. Les systèmes lotiques alimentés par des sources peuvent connaître de petites variations, principalement lorsqu'ils sont issus de sources souterraines qui prennent la température du sous-sol (Giller et Malqvist, 1998). De nombreux systèmes lotiques montrent d'importantes variations que ce soit du jour à la nuit ou de saison en saison. Ces variations sont plus ou moins importantes en fonction quelles se produisent en zones polaires, tempérées ou désertiques (Giller et Malqvist, 1998). L'exposition plus ou moins importante au soleil et l'altitude interviennent également et peuvent aussi influencer la température des systèmes lotiques (Allan, 1995).
Voir aussi
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « River ecosystem » (voir la liste des auteurs).
- Angelier 2003
- Allan 1995
- Giller et Malmqvist 1998
- Cushing and Allan 2001
- Brown 1987
Bibliographie
- (en) Allan, J.D. 1995. Stream Ecology: structure and function of running waters. Chapman and Hall, London. p. 388.
- (en) Angelier, E. 2003. Ecology of Streams and Rivers. Science Publishers, Inc., Enfield. p. 215.
- (en) Brown, A.L. 1987. Freshwater Ecology. Heinimann Educational Books, London. p. 163.
- (en) Cushing, C.E. and J.D. Allan. 2001. Streams: their ecology and life. Academic Press, San Diego. p. 366.
- (en) Edington, J.M., Edington, M.A., and J.A. Dorman. 1984. Habitat partitioning amongst hydrophyschid larvae of a Malaysian stream. Entomologica 30: 123-129.
- (en) Giller, S. and B. Malmqvist. 1998. The Biology of Streams and Rivers. Oxford University Press, Oxford. p. 296.
- (en) Hildrew, A.G. and P.S. Giller. 1994. Patchiness, species interactions and disturbance in the stream benthos. In Aquatic Ecology: scale pattern and process. (P.S. Giller, A.G. Hildrew, and D.G. Rafaelli eds.) Blackwell, Oxford. p. 21–62.
- (en) Morin, P.J. 1999. Community Ecology. Blackwell Science, Oxford. p. 424.
- (en) Rubbo, M. J. and J. M. Kiesecker. 2004. Leaf litter composition and community structure: translating regional species changes into local dynamics. Ecology 85:2519-2525.
- (en) Schleiter IM, Borchardt D, Wagner R, Dapper T, Schmidt Klaus-Dieter, Schmidt Hans-Heinrich, Werner Heinrich (1999), Modelling water quality, bioindication and population dynamics in lotic ecosystems using neural networks Ecological modeling 120 271–286 ; Elsevier
- (en) Schoener, T.W. 1974. Resource partitioning in ecological communities. Science 2:369-404.
- (en) Townsend, C.R., Hildrew, A.G., and K. Schofield. 1987. Persistence of stream invertebrate communities in relation to environmental variability. Animal Ecology 56:597-613.
- (en) Vannote, R.L., Minshall, G.W., Cummins, K.W., Sedell, J.R., and C.E. Cushing. 1980. The river continuum concept. Canadian journal of Fisheries and Aquatic Sciences 37:130- 137.
- (en) Vinson, M.R. and C.P. Hawknis. 1998. Biodiversity of stream insects: variation at local, basin, and regional scales. Annual Review of Entomology 43:271-293.
- (en) Ward, J.V. 1992. Aquatic Insect Ecology: biology and habitat. Wiley, New York. p. 456.
- (en) Watson, D.J. and E.K. Balon. 1984. Ecomorphological analysis of fish taxocenes in rainforest streams of northern Borneo. Journal of Fish Biology 25:371-384.
 Portail de l’écologie
Portail de l’écologie