Mycorhize éricoïde
La mycorhize éricoïde est une forme d'endomycorhize spécifique à la famille de plantes des Éricacées et de plusieurs champignons. Il s'agit d'une association obligatoire à bénéfices réciproques endotrophique, c'est-à-dire qu'elle se trouve à l'intérieur des cellules de la plante. Ce type d'association mycorhizienne est une des plus récemment apparues il y a 140 millions d'années[1].
.png.webp)
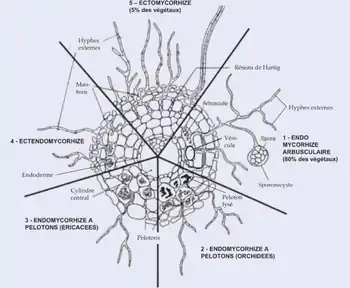
Description structurelle
Les Éricacées ont de fines racines blanches qui comportent d’une à trois couches de cellules : l'endoderme, l'hypoderme et l'épiderme. Ce sont les poils racinaires qui sont habituellement colonisés par l'endophyte. Les champignons ne pénètrent que la couche externe pour former des pelotons mycéliens qui remplissent les cellules. De plus, la membrane plasmique de chaque cellule s'invagine afin d'envelopper le mycélium. Habituellement, il n'y a qu'un seul point de pénétration par cellule, quoique plusieurs points d'entrée aient été observés à quelques reprises[2]. Il est important de noter que le champignon ne forme pas de manchon autour de la racine. Jusqu'à 80 % du volume de la racine peut être composé de tissus mycéliens[3].
Lors de l'extension de l'apex racinaire, de nouvelles cellules épidermiques sont différenciées. L'hyphe, qui ne forme qu'un réseau autour des zones de la radicelle ayant un épiderme mature croît lors de cette extension. Il est rare qu'il se rende jusqu'à l'apex racinaire, quoique cela puisse se produire dans des conditions où la croissance de la racine est particulièrement lente[4].
Échanges de nutriments

L'humus qui supporte la croissance des plantes Éricacées étant pauvre, acide et peu minéralisé, le champignon éricoïde symbiote joue un rôle important dans l'acquisition de nutriments à travers la mycorhize éricoïde. Il possède plusieurs caractéristiques saprotrophiques qui lui permettent de dégrader la matière organique pour ensuite la redistribuer. En outre, il produit plusieurs enzymes hydrolytiques et oxydatives qui aident à la mobilisation de molécules organiques présentes sous forme récalcitrante dans le sol. Ces enzymes ont pour la plupart une activité optimale à de faibles pH. Par exemple, les sols des forêts boréales comportent de fortes concentrations de parois de champignons mal décomposées, la chitine, qui contient environ 40 % d'azote. Or un champignon éricoïde comme Pezoloma eriae est capable de la dégrader et de donner accès à cette ressource aux Ericacées symbiotes comme par exemple un myrtillier. Le phosphore est aussi rendu accessible à la plante grâce à la digestion de sa forme organique par le champignon éricoïde[5],[6],[7].
En plus d'accéder à des bénéfices nutritifs par cette association, les Éricacées profitent d'avantages protecteurs. En effet, les champignons symbiotes ont la capacité de séquestrer des ions métalliques qui sont toxiques pour la plante. Ceci empêche les toxines d'accéder aux pousses de la plante et d'interférer avec la photosynthèse[8].
De leurs côtés, comme dans toutes associations mycorhiziennes, les champignons profitent de l'apport de sucres, de lipides et de vitamines de la part des plantes qui les produisent grâce à la photosynthèse[6].
Habitat
Les sols dans lesquels poussent les Éricacées et les champignons éricoïdes sont caractérisés par une faible minéralisation de la matière organique, de faibles teneurs en nutriments comme l'azote, le phosphore et le potassium, une forte acidité et une quantité importante de tanins. Leur humus est de type Mor[1],[6]. La litière produite par les Éricacées est elle-même peu minéralisable, ce qui a pour conséquence d'éliminer la concurrence des plantes en incapacité de vivre dans ces milieux car dépourvue de mycorhizes éricoïdes[6].
Leurs biotopes sont la taïga, la lande et la toundra sur tous les continents sauf en Arctique[1],[6].
Étant donnée leur vaste distribution géographique, ces plantes vivent dans une grande diversité de conditions hydrologiques. Ceci provoque différents cycles de croissance des racines des Éricacées, ainsi que des cycles de colonisation par les champignons éricoïdes très variés[9]
Espèces de champignons concernées
_1.jpg.webp)
_Tul._%2526_C._Tul_292965.jpg.webp)
Historiquement, les recherches sur la physiologie et la fonction des champignons mycorhiziens éricoïdes se sont concentrées sur des isolats fongiques morphologiquement identifiés comme l'ascomycète Pezoloma ericae (anciennement connu sous le synonyme Rhizoscyphus ericae)[10]. D'autres espèces d'ascomycètes forment également des mycorhizes éricoïdes comme le genre Meliniomyces et les espèces Cairneyella variabilis, Gamarada debralockiae et Oidiodendron maius[11],[12],[13].
L'application du séquençage de l'ADN à des isolats fongiques a permis de découvrir une grande diversité de communautés fongiques dans les racines d'Éricacées. Cependant, la capacité de ces champignons à former des enroulements mycorhiziens éricoïdes typiques n'a pas été vérifiée et certains pourraient être des endophytes, des saprobiontes ou des parasites non mycorhiziens[14],[15],[16],[17].
Chez les basidiomycètes, les espèces de l'ordre des Sebacinales comme Sebacina incrustans sont des associés fréquents qui peuvent former des mycorhizes éricoïdes[14],[15],[18], de même qu'une seule espèce de l'ordre des Hymenochaetales, Kurtia argillacea[19].
Notes et références
- Morrissette, N. (2011). Les mycorhizes éricoïdes : un potentiel biotechnologique pour favoriser l'établissement de plants de bleuet sur les sites perturbés par l'exploitation des sables bitumineux en forêt boréale canadienne, M. Sc. Université Laval.
- Smith, S., Read, D. and Harley, J. (1997), Mycorrhizal symbiosis, San Diego, Calif.: Academic Press.
- Durrieu, G. (1993), Écologie des champignons, Paris: Masson. p. 71.
- Smith, S., Read, D. and Harley, J. (1997), Mycorrhizal symbiosis, San Diego, Calif.: Academic Press. p. 326.
- Morrissette, N. (2011), Les mycorhizes éricoïdes : un potentiel biotechnologique pour favoriser l'établissement de plants de bleuet sur les sites perturbés par l'exploitation des sables bitumineux en forêt boréale canadienne, M. Sc. Université Laval
- Marc-André Selosse, L’origine du monde : Une histoire naturelle du sol à l’intention de ceux qui le piétinent, Arles, Actes Sud, , 480 p. (ISBN 978-2330152673)
- (en) Read DJ , Leake JR , Perez-Moreno J., « Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes », Canadian Journal of Botany, vol. 82, , p. 1243–1263 (DOI 10.1139/b04-123, lire en ligne)
- Smith, S., Read, D. and Harley, J. (1997), Mycorrhizal symbiosis, San Diego, Calif.: Academic Press. p. 346.
- Read, D. (1996), The structure and function of the ericoid mycorrhizal root, Annals of Botany, 77(4), p. 365-374
- Smith, S. E. and D. J. Read. 2008. Mycorrhizal Symbiosis, Third Edition. Academic Press
- Hambleton S, Sigler L (2005) Meliniomyces, a new anamorph genus for root-associated fungi with phylogenetic affinities to Rhizoscyphus ericae (≡ Hymenoscyphus ericae), Leotiomycetes. Studies in Mycology. 53:1-27.
- Midgley, D.J., Rosewarne, C.P., Greenfield, P., Li, D., Vockler, C.J., Hitchcock, C.J., Sawyer, N.A., Brett, R., Edwards, J., Pitt, J.I. & Tran-Dinh, N. (2016). Genomic insights into the carbohydrate catabolism of Cairneyella variabilis gen. nov., sp. nov., the first reports from a genome of an ericoid mycorrhizal fungus. Mycorrhiza, 26: 345–352.
- Midgley, D.J., Sutcliffe B, Greenfield P & Tran-Dinh, N. (2018) Gamarada debralockiae gen. nov. sp. nov.—the genome of the most widespread Australian ericoid mycorrhizal fungus. Mycorhiza, 28: 379-389.
- Allen, T. R., T. Millar, S. M. Berch, and M. L. Berbee. 2003. Culturing and direct DNA extraction find different fungi from the same ericoid mycorrhizal roots. New Phytologist 160:255-272.
- Selosse, M. A., S. Setaro, F. Glatard, F. Richard, C. Urcelay, and M. Weiss. 2007. Sebacinales are common mycorrhizal associates of Ericaceae. New Phytologist 174:864-878.
- Kjoller, R., M. Olsrud, and A. Michelsen. 2010. Co-existing ericaceous plant species in a subarctic mire community share fungal root endophytes. Fungal Ecology 3:205-214.
- Walker, J. F., L. Aldrich-Wolfe, A. Riffel, H. Barbare, N. B. Simpson, J. Trowbridge, and A. Jumpponen. 2011. Diverse Helotiales associated with the roots of three species of Arctic Ericaceae provide no evidence for host specificity. New Phytologist 191: 515-527.
- Vohník M, Pánek M, Fehrer J, Selosse M-A (2016) Experimental evidence of ericoid mycorrhizal potential within Serendipitaceae (Sebacinales). Mycorrhiza 26:831–846
- Kolarik M, Vohnik M (2018) When the ribosomal DNA does not tell the truth: the case of the taxonomic position of Kurtia argillacea, an ericoid mycorrhizal fungus residing among Hymenochaetales. Fungal Biology 122:1–18
 Portail de la botanique
Portail de la botanique  Portail de la mycologie
Portail de la mycologie