Hermaphrodisme successif
L'hermaphrodisme successif (ou séquentiel) est une forme d'hermaphrodisme correspondant à une acquisition des caractères sexuels mâles et femelles différée dans le temps. Il se retrouve aussi bien chez les animaux que chez les plantes, mais pour ces dernières on préférera le terme dichogamie.


Contrairement à l’hermaphrodisme simultané, la production des gamètes mâles et femelles s’effectue à deux temps distincts. Ce mode de reproduction peut être considéré comme un intermédiaire entre le gonochorisme et l'hermaphrodisme simultané, ces derniers étant les extrêmes d'un même continuum[1]. Par ailleurs, l'hermaphrodisme simultané n'est pas nécessairement constant tout au long de la vie d'un même individu. Il peut être précédé par une phase d'hermaphrodisme successif. Dans ce cas, chaque individu passe ainsi successivement par trois stades[2].
On peut distinguer trois grandes tendances d’hermaphrodisme séquentiel : la protandrie (ou protérandrie) où l’individu est d’abord mâle puis devient femelle ; la protogynie (ou protérogynie) où l’individu est d’abord femelle puis devient mâle ; et l’hermaphrodisme alternant où l’individu va changer de sexe plus d’une fois au cours de sa vie[1].
Hypothèse d'apparition
La répartition de l’hermaphrodisme dans le règne animal est encore mal comprise, néanmoins certaines hypothèses se proposent d’expliquer l’apparition des différents types de sexualités au sein de certains clades. Une étude portant sur les crustacés suggère que l’hermaphrodisme séquentiel serait un hermaphrodisme secondaire issu du gonochorisme[3]. Dans cette hypothèse, l’hermaphrodisme simultané est considéré comme le type de sexualité ancestrale dont est issu le gonochorisme[3]. Des conflits génétiques liés à la détermination du sexe seraient ensuite apparus chez des individus gonochoriques, causés par des mutations ou des insertions de séquences parasites influant sur ces gènes de déterminations et créant un délai dans l’expression de ceux-ci, d’où la succession chronologique des phases sexuelles chez les hermaphrodites séquentiels[3].
L'hermaphrodisme est la forme de mise en place des appareils reproducteurs la plus répandue chez les plantes à fleurs (Angiospermes). La dichogamie, qui correspond à la forme successive de l'hermaphrodisme, fait partie des traits évolutifs floraux mis en place par les Angiospermes pour faire face à la cohabitation des fonctions mâles et femelles au sein d'une même fleur. Chez les angiospermes, bien que la fréquence exacte de la dichogamie soit inconnue, des observations suggèrent une haute fréquence (87% d'espèces recensées sur 4277). Et bien que la fonction significative de la dichogamie reste encore incertaine, son rôle est toutefois reconnu comme important dans la biologie florale (Bertin and Newman, 1993) . De nombreuses hypothèses sont posées, notamment l'implication de la dichogame dans la réduction de la dépression de consanguinité et la réduction de l'autofécondation, conséquence de la géinotogamie (pollinisation par le pollen d'autres fleurs de la même plante)[5].
Sélection sur la fitness
Avantage de la taille
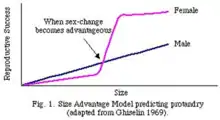
Chez les espèces animales hermaphrodites séquentielles, le changement de sexe a été expliqué par la différence de succès reproducteur (ou fitness) entre le sexe mâle et femelle pour une même taille ou un même âge donné(e) : c’est le size-advantage model proposé par Ghiselin (1969)[1]. Si cette différence est significative au sein d’une espèce, la pression de sélection va alors s’exercer en faveur de l’hermaphrodisme séquentiel, car il sera plus avantageux pour un individu d’une certaine taille d’être d’un certain sexe à un moment donné. Plus l’allocation sexuelle sera grande pour un sexe, plus un individu de ce sexe aura intérêt à être gros pour s’assurer une fécondité importante. Il est donc opportun pour un individu de changer de sexe lorsque sa fitness pour la taille atteinte est inférieure à la fitness qu’il aurait en étant du sexe opposé[1]. La théorie des constantes sans dimensions proposé par Charnov et Skuladottir permet de prédire, à partir de paramètres connus, à quel moment (âge ou taille) le changement de sexe devrait intervenir chez un individu hermaphrodite[6].Cette théorie a été testée empiriquement sur 52 espèces de poissons protogynes et protandres, et les résultats ont permis d’estimer que le changement de sexe intervenait pour une taille de 80% de la taille maximum et pour un âge correspondant à 2,5 fois l’âge de la maturité[6].
En parallèle chez les végétaux la dichogamie a été un mécanisme initialement décrit comme permettant d'éviter l'autofécondation. Cependant, ce mécanisme semblerait être un trait floral sélectionné pour d'autres raisons. En effet la réduction de l'interférence sexuelle (causé par la dichogamie) favorise le croisement chez les plantes auto-compatibles (diminuant l'autofécondation) et pourtant ce trait se retrouve aussi chez les auto-incompatibles (absence d'autofécondité) (Bertin, 1993). La conservation de ce trait, notamment chez les auto-incompatibles, pourrait être expliqué par la morphologie des angiospermes. En effet il semblerait que la réduction de l'interférence sexuelle entraine une meilleure dispersion du pollen et un meilleur développement de la taille des pistils, en diminuant la gêne physique et en permettant une meilleure allocation à la fonction sexuelle présente et ainsi augmenter la fitness de l'individu [7]. Soit plutôt comme un mécanisme général pour réduire l’impact de l’interférence pollen-pistil sur l’import et l’export du pollen (Lloyd & Webb, 1986; Barrett, 2002). Contrairement à l’hypothèse de l’évitement de la consanguinité, qui se focalisait sur la fonction femelle. Cette hypothèse d’interférence-évitement considère les fonctions des deux genres, et a été confirmée par un modèle de l'évolution de la dichogamie incluant la sélection sex-ratio, l'interférence anthère-stigma et la dépression de consanguinité, par la conservation de ce trait floral chez les auto-incompatibles dans l'évolution de la dichogamie[8].
Évitement de la dépression de consanguinité/export gamètes mâles
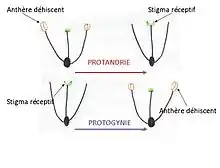
La dépression de consanguinité chez les végétaux intervient lorsque l'autofécondation devient désavantageuse, c’est-à-dire lorsque les descendants d'autofécondation possèdent une fitness moindre que celle de descendants croisés. Ceci est le plus souvent causé par des gènes délétères récessifs qui se retrouvent combinés chez les descendants homozygotes des consanguins. C'est pour cela que la dichogamie semblerait conférer un certain avantage à la fitness, des auto-compatible, en évitant ce phénomène par le mécanisme sexuel qu'est la dichogamie. Cependant le rôle exact de la dépression de consanguinité dans l'évolution de la dichogamie reste incertain[8].
Sélection sur la direction du changement de sexe
Protandrie favorisée

La qualité de la gamétogenèse chez la plupart des animaux mâles est peu variable selon leur taille. À l'inverse, la gamétogenèse chez les femelles est souvent plus coûteuse en énergie ce qui induit une corrélation avec la taille sur le plan de la qualité (taille des gamètes plus importante)[9]. Un mâle de petite taille pourra donc féconder un grand nombre de femelles. Cependant, le succès reproducteur d’une femelle de petite taille sera moindre car sa fécondité est dépendante de sa taille.
Dans les cas où cette différence d’allocation sexuelle est très importante, la protandrie sera favorisée et les individus les plus gros (et donc les plus vieux) maximiseront leur fitness en étant de sexe féminin. Les observations effectuées à ce jour suggèrent que la protandrie est le mode d’hermaphrodisme séquentiel le plus répandu dans le règne animal[3]. On le retrouve chez un grand nombre d’espèces de mollusques (exemple de la crépidule).
Chez les angiospermes, des études suggèrent que le mode de pollinisation aurait un rôle fonctionnel dans l'évolution du type de dichogamie (Bertin and Newman, 1993). En effet, les espèces protandres semblent avoir tendance à être pollinisées par des abeilles et/ou des mouches, correspondant à des facteurs biotiques. De plus, du fait de la durée de viabilité du pollen dans de nombreuses espèces, la protandrie ne semblerait pas être efficace comme adaptation pour éviter l'autopollinisation. Elle serait plutôt associée à une architecture d'inflorescence particulière ainsi qu'un comportement de pollinisateurs pour réduire l'autofécondation et la réduction du pollen disponible pour la dispersion[10].
Protogynie favorisée
Chez les animaux, la protogynie sera favorisée principalement dans les systèmes sociaux régis par un mâle dominant. Seuls les plus gros individus peuvent avoir un succès reproducteur important en tant que mâle. Les plus petits ont donc intérêt à être femelle pour maximiser leurs fitness et éviter la compétition. On peut observer ce système de reproduction chez beaucoup d’espèces de poissons de récifs coralliens. Lorsque le mâle dominant contrôlant le harem meurt, la femelle de plus grande taille change de sexe et prend sa place[1]. Elle est très répandue dans certaines familles de téléostéens.
La protogynie est supposée être un trait ancestral chez les angiospermes (Endress, 1997). Elle aurait tendance à être groupée dans les familles à la base du branchement des angiospermes. Ce trait semble être un moyen pour prévenir l'autofécondation (Lloyd and Webb, 1986). En effet, les ovules peuvent être fécondés par du pollen de sources externes, avant que la fleur ne présente son pollen. De plus, s'il reste des ovules non fécondés, ils peuvent l'être par autofécondation, lors de l'expulsion du pollen. Enfin, les plantes à fleurs protogynes sembleraient être préférentiellement pollinisé par le vent, facteur abiotique, hypothèse confirmée par le modèle de Pagel (1994) [10].
Alternant favorisé
Chez les angiospermes, ce type d’hermaphrodisme est assez rare[1]. Arisaema triphyllum, par exemple, peut être soit mâle soit femelle au cours d’une saison de reproduction, son changement de sexe s’effectuant en réponse à l’environnement d’une saison sur l’autre.
Chez les animaux, l’hermaphrodisme alternant peut être accommodant pour la reproduction d’une population : chez certain poisson, le changement de sexe d’un individu va permettre de reformer une paire fonctionnelle dans le cas de la perte d’un partenaire ou d’assurer la reproduction lorsque la recherche de partenaire sexuel est trop dangereuse[1]. Un autre exemple peut être observé chez l’huître, dont la capacité reproductive en tant que femelle est limitée par la taille de la chambre de gestation et non pas par l’énergie disponible ; cette énergie en excès va être utilisée pour la production de sperme durant la période de « gestation » des œufs fertilisés[1]. Ainsi, l’huître maximise sa fitness en étant successivement femelle (production d'œufs) puis mâle (spermatogenèse) au cours de son cycle de reproduction.
Facteurs du changement de sexe
Génétiques
D’un point de vue génétique, le changement de sexe chez les animaux est le plus souvent lié à l’âge ou à la taille de l’individu [11]. Les différences en croissance et en taux de mortalité des sexes jouent également un rôle moteur dans l'hermaphrodisme séquentiel. Si un sexe a un taux de mortalité plus élevé, il est plus avantageux d'être du sexe opposé. De la même manière, si un sexe a un taux de croissance plus élevé, il sera plus avantageux d'être de ce sexe en premier lieu.
Abiotiques
Il y a des avantages évolutifs à être un hermaphrodite séquentiel. La capacité à changer de genre en réponse aux pressions environnementales fait des hermaphrodites séquentiels des individus bien plus flexibles. Ils sont ainsi susceptibles de faire face à des situations qui pourraient conduire à une baisse de population voire à une extinction pour d’autres espèces. Différents paramètres environnementaux peuvent influer, comme la température, les ressources en nourriture, la salinité du milieu, la pollution … L’environnement peut ainsi influer sur le sex-ratio. C’est le déterminisme environnemental du sexe (ESD)[11]. Par exemple, les peintures anti fouling, utilisées par les coques des bateaux, contiennent de l’étain-tributyle qui provoque le développement de caractères mâles chez les femelles, source de stérilité chez certains gastéropodes (pseudo-hermaphrodisme).
Sociaux
On trouve ces facteurs du changement de sexe chez une multitude d’espèces animales, essentiellement marines. Ces facteurs sociaux sont principalement dus à des problématiques de sex-ratio au sein d'une population et peuvent ainsi se manifester dans des situations de déséquilibre des populations [11]. C’est le cas de Thalassoma bifasciatum, un petit poisson qui vit territorialement au niveau de récifs. Selon la taille du récif, un ou plusieurs mâles peuvent être présents pour un harem de femelles. Cette espèce se caractérise par un hermaphrodisme séquentiel de type protogynie. Au sein de ce harem, un ou plusieurs mâles (selon la taille du récif) seront dominants. Ainsi pour les autres individus de la population, la stratégie optimale consiste à commencer le cycle de vie comme un individu de type femelle. Quand un mâle territorial meurt, la plus grande femelle change de sexe et obtient ainsi le territoire qui s'accompagne du statut de mâle dominant. Les femelles changent ainsi de sexe en réponse à un manque de mâles à proximité. En effet, à la mort du mâle dominant, des hormones de stress sont sécrétées induisant un changement de sexe pour la femelle dominante.
Notes et références
- (en) Janet L. Leonard, « Williams' Paradox and the Role of Phenotypic Plasticity in Sexual Systems », sur Integrative and Comparative Biology,
- Micheline Martoja, Mollusques, Institut océanographique, , p. 95
- (en) Pierre Juchault, « Hermaphroditism and gonochorism. A new hypothesis on the evolution of sexuality in crustacea », sur Comptes rendus de l'Académie des sciences,
- (en) Eduardo Narbona, Pedro L. Ortiz, Montserrat Arista, « Linking Self-Incompatibility, Dichogamy, and Flowering Synchrony in Two Euphorbia Species: Alternative Mechanisms for Avoiding Self-Fertilization? », sur PLOS one,
- (en) D.J. Allsop & S.A. West, « Constant relative age and size at sex change for sequentially hermaphroditic fish », sur Journal of Evolutionary Biology,
- (en) Can Dai,Laura F. Galloway, « Do dichogamy and herkogamy reduce sexual interference in a self-incompatible species? », sur Functional Ecology,
- (en) Risa D. Sargent, Mohammad A. Mandegar and Sarah P. Otto, « A model of the evolution of dichogamy incorporating sex ratio selection anther-stigma interference and inbreeding depression », sur Evolution,
- (en) Risa D. Sargent and Sarah P. Otto, « A phylogenetic analysis of pollination mode and the evolution of dichogamy in angiosperms », sur Evolutionary Ecology Research,
- (en) Robert R. Warner, « Sex change and the size-advantage model », sur Trends in Ecology and Evolution,
Voir aussi
 Portail de la biologie
Portail de la biologie  Portail de la botanique
Portail de la botanique  Portail de la zoologie
Portail de la zoologie